Ключевые выводы:
Удаление всех вторичных ветвей от основных в течение вегетационного периода лучше всего повысило химическую однородность соцветий.
Наивысшего урожая каннабиноидов удалось достичь за счёт увеличения урожайности соцветий при двойной обрезке — в день пересадки черенков в горшки и при смене фотопериода на цветение.
Манипуляции с формой растения могут использоваться для увеличения урожайности соцветий, но увеличения концентрации каннабиноидов следует достигать иными способами.
Примечание: Закономерности были обнаружены при исследовании КБД-доминантного сорта «Topaz» от BOL Pharma.
Вступление
Постоянный интерес к каннабису обусловлен воздействием многочисленных биологически активных вторичных метаболитов (включая, терпены, флавоноиды и уникально продуцируемые каннабиноиды), обнаруженных в основном в соцветиях. Помимо известных психоактивных эффектов, каннабис полезен для лечения многих заболеваний, включая неврологические состояния, обезболивание и многое другое. Терапевтические эффекты объясняются биологическим взаимодействием между комбинациями вторичных метаболитов и рецепторами эндоканнабиноидной системы. Изменения химического профиля потребляемого растительного материала, который очень разнообразен и включает широкий спектр хемотипов, может привести к изменению эффективности.
Хотя потенциал производства определённого профиля вторичных метаболитов в каннабисе определяется генетическим фоном растения, на фактические уровни продуцируемых метаболитов в значительной степени влияют условия окружающей среды во время культивирования. К ним относятся:
Минеральное питание.
Интенсивность и спектр света.
Стрессовые условия.
Изменчивость химического профиля между соцветиями наблюдается по всей длине растения. Более того, поскольку органы растений локально воспринимают окружающую среду, различия между микроклиматом внутри побегов дополнительно вызывают изменения в физиологии и вторичном метаболизме. Чтобы повысить однородность химического профиля внутри одного растения, между разными растениями и между циклами выращивания, важно понимать, как различные микроклиматические условия и методы культивирования влияют на вторичный метаболизм. Это позволит использовать новые методы выращивания для смягчения химических вариаций в растениях за счёт минимизации микроклиматических градиентов. Настоящее исследование было направлено на использование манипуляций с архитектурой растений для стандартизации профиля каннабиноидов у крупных медицинских растений каннабиса.
Архитектура растений оказывает огромное влияние на микроклимат побегов, влияя на проникновение света, влажность и температуру. В системах сельскохозяйственного производства используются несколько методов для изменения архитектуры растений, включая обрезку основного стебля/ветвей, удаление ветвей и шпалеровку. Ещё один способ воздействия на климат в пологе, не влияющий на структуру растения, — полное или частичное удаление листьев. Сообщается, что изменение климата в побегах вызывает изменения качества урожая, такие как повышение питательной ценности из-за дефолиации у бобовых и винограда, а также качества у обрезанных дынь и болгарского перца. Более того, такие архитектурные изменения могут как увеличить, так и уменьшить количество урожая.
Что касается каннабиса, то в одном исследовании с промышленными сортами конопли проверялись эффекты обрезки основного стебля и сообщалось о повышении урожайности семян. Сообщалось, что у каннабиса «лекарственного типа» удаление ветвей снижает биомассу урожая и вызывает изменения в химическом профиле, который варьируется между сортами и каннабиноидами. Для многих видов растений документально подтверждено, что изменения в архитектуре растений влекут за собой влияние на интенсивность и спектр света внутри полога. Было обнаружено, что у каннабиса изменения интенсивности и спектра света влияют на количество и качество урожая, предполагая потенциал архитектурных манипуляций растений для регуляции локализованного вторичного метаболизма и пространственной стандартизации.
Пространственные градиенты интенсивности света в пологах растений усиливаются с высотой растения, поскольку доля света, достигающего нижних частей растения, уменьшается с увеличением продольного расстояния от источника света. Следовательно, вероятность вызванных микроклиматом изменений физиологических и химических свойств вдоль растений выше у растений с высоким пологом, чем у более мелких растений. Поэтому обработки, регулирующие архитектуру растений, имеют потенциал для смягчения этих эффектов за счет увеличения проникновения света в побеги. В индустрии производства каннабиса методы варьируются от выращивания небольших коротких растений в контролируемом «индоре» до более крупных растений в теплицах или в «аутдоре». Значительная часть товарного производства основана на интенсивном выращивании крупных растений. Пространственные градиенты химических свойств и химической однородности у крупных растений каннабиса ранее не изучались.
Материалы и методы
Растительный материал и условия выращивания
Для исследования использовали медицинский сорт каннабиса «Topaz» (BOL Pharma, Ревадим, Израиль). Это сорт типа III, содержащий высокий уровень КБД (8–16%) и низкий уровень ТГК (<1%). Эксперимент проводился на сертифицированной коммерческой ферме по выращиванию каннабиса в теплице с естественным освещением с добавлением фотопериодического света. Растения были выведены из черенков в кокосовом субстрате. Укорененные черенки высаживали в горшки объёмом 13 л, по 1 растению на горшок, в смесь торфяных мхов, с плотностью 1 растение/м2.
Растения случайным образом делили на восемь групп по шесть растений в каждой. На стадии вегетативного роста растения культивировали при длительном фотопериоде 24/0 (свет / темнота) часа, а фотопериодическое освещение дополняли люминесцентными лампами.
После 63 дней вегетативного роста растения были переведены на короткий фотопериод продолжительностью 12 часов, чтобы вызвать цветение. Удобрения подавались путем фертигации, т.е. растворялись в ирригационном растворе. Орошение проводилось через капельницы (по 4 на каждый горшок) с регулируемым расходом 1,2 л/ч. Объём поливной воды при каждом поливе был установлен так, чтобы обеспечить дренаж примерно в 30%, и он увеличивался на протяжении всего развития растений до 3 л /день с каждого горшка. Опыт был прекращён через 111 дней после посадки или через 58 дней после перехода к короткому световому периоду на стадии созревания, принятой для коммерческого сбора.
Способы обработки растений
На каждой группе были оценены по одному из восьми способов обработки каннабиса:
Контрольная группа, которая не подвергалась обработке (Control)
Дефолиация за 3 недели до сбора урожая (Defoliation)
Удаление ветвей и листьев с нижней (1/3) части растения при переходе на короткий фотопериод (BLBR)
Удаление ветвей и листьев с нижней части растения при переходе на короткий фотопериод + Дефолиация (BLBR + Defoliation)
Удаление всех ветвей от главного стебля в течение вегетационного периода (1° Branch removal)
Удаление всех вторичных ветвей от основных ветвей в течение вегетационного периода (2° Branch removal)
Обрезка укоренившихся черенков в день пересадки с оставлением шести основных ветвей (Single prune)
Обрезка укоренившихся черенков в день пересадки с оставлением шести основных ветвей + обрезка при переходе на короткий фотопериод (Double prune)
Рост растений, накопление биомассы, PAR и урожайность
Каждые две недели измеряли высоту каждого растения от основания растения до вершины апикальной меристемы главного стебля (при обработке обрезкой измеряли высоту самой высокой ветви).
При окончательном сборе урожая биомассу соцветий, стеблей и веерных листьев измеряли для каждого растения индивидуально с помощью промышленной шкалы «Mierav 4000». Затем соцветия обрезали промышленным триммером для удаления выступающих соцветий. Листья и обрезанные соцветия снова взвешивались для расчёта биомассы обрезанных листьев соцветий.
Урожайность соцветий оценивалась после сушки в соответствии с общепринятым отраслевым стандартом 15% воды в ткани.
Урожайность каннабиноидов рассчитывалась путём умножения средней концентрации каждого каннабиноида в растении на биомассу выхода сухого веса соцветий растения.
Активное фотосинтетическое излучение (PAR) измеряли в четырёх точках на каждом участке на четырёх высотах вдоль растения (0, 0,5, 1,2 и 2 м от основания) с помощью квантового датчика Apogee MQ-500.
Результаты
Развитие полога
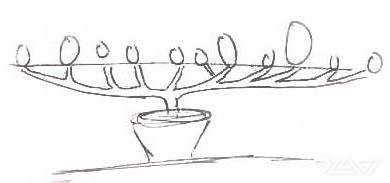
Схема роста и форма побегов растений обычно регулируются цветоводами во всём мире с помощью множества методов, включая густоту посадки, ScrOG, гормоны растений и физическое удаление органов растений. На рисунке ниже изображена реакция крупных растений каннабиса на семь воздействующих на архитектуру обработок по сравнению с контрольным растением:
Рис. 1: Реакция каннабиса на тренировки
Структура растения не изменилась визуально при дефолиации и удалении всех вторичных ветвей в течении вегетационного периода, а также при обеих обработках «BBLR» только нижняя часть полога отличалась по форме от контрольного растения.
Растения после двойной обрезки (Double prune) имели форму, аналогичную контрольным растениям, но имели меньший рост.
После удаления всех ветвей от главного стебля в течение вегетационного периода (1° Branch removal) растение состояло из одного удлинённого прямого стебля без ветвей. Одинарная обрезка (Single prune) вызвала развитие двух стеблевидных ветвей.
Высота растений варьировалась между обработками. Уже через 7 дней после начала обработки было измерено статистически значимое уменьшение высоты обрезанных растений. Примерно через 67 дней после начала обработок, то есть через 7 дней после перехода на режим короткого дня и второй обрезки, растения «одинарной обрезки» вытянулись в высоту, компенсируя уменьшение высоты, вызванное обрезкой, и растения достигли высоты контрольных растений.
Архитектурные манипуляции, которые включают ранение и удаление органов растений, изменяют эндогенные программы развития, влияя на апикальное доминирование и, следовательно, также на гормональные профили. Выявленные у растений три изменения в развитии указывают на изменение гормональной активности.
Во-первых, две полуглавные ветви, полученные при обработке «одинарной обрезкой», можно объяснить повышенным количеством гибберелловой кислоты, которая стимулирует как удлинение растений, так и ингибирование развития боковых почек. Такое увеличение выработки активного гиббереллина ранее наблюдалось у различных многолетних растений и может также объяснить, почему при этой обработке у растений не происходит уменьшения высоты.
Во-вторых, вторая обрезка была осуществлена при переходе к режиму короткого дня, когда компактные сложные соцветия каннабиса начинают развиваться за счёт ограничения удлинения ветвей и развития коротких веточек. Возможно, этот сдвиг в развитии связан с катаболизмом гиббереллина, что также могло бы объяснить более короткие растения при обработке «двойной обрезкой» и усиление ветвления, которое приводит к развитию более «густых» растений при этой обработке. Отсутствие индуцированного гиббереллином покоя почек при переключении на короткий фотопериод может также объяснить стимуляцию роста и развития пазушных почек по всему растению.
В-третьих, удаление всех ветвей от главного стебля в течение вегетационного периода вызвало удлинение основного стебля больше, чем при всех других процедурах. Когда эта процедура применялась к более мелким медицинским растениям каннабиса, такой же фенотипический ответ был обнаружен у одного из двух изученных генотипов.
Цитокинин — это фитогормон, который вызывает деление клеток и тесно связан с ветвлением растений. Молекулы, полученные из корней, транспортируются в побеге акропетально (от основания к вершине), вызывая развитие и ветвление пазушных почек. Удаление ветвей с растения приводит к более высокому отложению цитокинина в апикальной меристеме, вызывая повышенную активность меристемы и удлинение растения.
Когда свет проходит через полог растения, он поглощается как листьями, так и ветвями, изменяя как интенсивность, так и спектр. Интенсивность света была измерена на четырёх различных высотах, а результаты представлены на рисунке ниже:
Рис. 2: А — Время от начала вегетативного роста; В — Высота растения
Интенсивность постепенно снижается с уменьшением высоты по направлению к нижней части растений. Степень уменьшения и уровень интенсивности по вертикальному профилю различались в зависимости от способа обработки. Наибольшая интенсивность у основания растения была получена для обеих дефолиаций, что демонстрирует потенциал дефолиации для уменьшения градиентов микроклимата.
Обе обработки «BBLR» имели более высокую интенсивность света внизу растения, чем на высоте 50 см над землёй. Вероятно, это вызвано горизонтальным проникновением света в эту часть растений.
Интенсивность света на высоте 50 см над землёй была одинаковой для обеих дефолиаций. Это указывает на то, что свет, поглощаемый ветвями и соцветиями, был одинаковым в обеих обработках, и что, несмотря на удаление нижних ветвей, плотность побегов была одинаковой в обеих обработках, что также было продемонстрировано воздействием обработок на биомассу растений. Разница в интенсивности света у основания растения между обработками «BBLR + дефолиация» и «дефолиация» была аналогична разнице между обработками «BBLR» и «Контроль» и была небольшой в обоих случаях — от 25 до 41 мкмоль/м2*с.
Проникновение света в побеги во многом влияет на развитие растений. Во-первых, повышенная интенсивность света в нижней части побега идёт рука об руку с более тёплым и сухим микроклиматом. Повышенная освещённость и пониженная относительная влажность способствуют скорости транспирации и фотосинтеза, поддерживая ускоренный рост. Это было продемонстрировано во многих системах земледелия. Например, в баклажане (Solanum melongena) повышенный фотосинтез был зарегистрирован у растений, обрезанных для увеличения проникновения света в полог, что привело к увеличению содержания ассимилятов углерода. Такое улучшенное функционирование растений желательно в интенсивном сельском хозяйстве, и поэтому манипуляции с растительным покровом часто используются для увеличения проникновения света. У камелии масличной (Camellia oleifera) форма с открытым центром увеличила проникновение света и температуру в нижней части полога, а также снизила относительную влажность. Этот изменённый микроклимат увеличил урожай семян и содержание масла в нижней части полога по сравнению с круглой закрытой формой полога. У яблонь проникновение света положительно коррелирует с плотностью цветочных бутонов, урожайностью плодов, цветом кожуры плодов, содержанием растворимых твердых веществ и плотностью плодов.
Учитывая возможность улучшения химического состава за счет увеличения проникновения света, следующим был исследован профиль каннабиноидов растений.
Химический ответ
Разница между выращиванием крупных и мелких растений влечёт за собой различия в физиологии растений и большие различия в микроклиматических условиях вдоль всего растения. Более длинное расстояние между верхушкой побега и основанием растения влечёт за собой большие градиенты гормонов и микросреды и более низкие значения у основания растения. Кроме того, у более высоких растений большее количество органов (в центре и внизу растений) страдает от затенения, так как больше листьев и ветвей выше поглощают свет.
Поскольку на развитие соцветий каннабиса влияют как эндогенные, так и экзогенные факторы, вполне вероятно, что более крупные растения будут страдать от повышенной пространственной изменчивости по сравнению с более мелкими растениями. Чтобы сравнить вариации вдоль растения и между обработками, среднюю концентрацию каждого каннабиноида в каждом месте отбора пробы сравнивали с концентрацией в первичном апикальном соцветии (точка I) контрольного растения. На рисунке ниже — чем дальше точка данных находится от центра, тем выше концентрация каннабиноида в конкретном месте по сравнению с концентрацией контрольных растений:
Относительные концентрации шести каннабиноидов, на которые влияют обработки с изменением архитектуры
В основном пути биосинтеза каннабиноидов каннабигероловая кислота (КБГ-К) является первым образовавшимся каннабиноидом, который служит предшественником ферментативно-катализируемого биосинтеза первичных каннабиноидов ∆9 -тетрагидроканнабиноловой кислоты (ТГК-К) и каннабидиоловой кислоты (КБД-К).
Концентрация КБГ-К продемонстрировала зависимый от метода обработки растения ответ. Самый высокий уровень был у растений при удалении всех ветвей от главного стебля в течение вегетационного периода (1° Branch removal). У этих растений концентрация КБГ-К достигла уровня в 3,7–4,6 раза выше, чем у контрольных растений.
Поскольку КБГ-К является предшественником всех других каннабиноидов, его концентрация в растении является динамичной и отражает чистую активность его биосинтеза и дальнейшую трансформацию по каннабиноидным путям. Возникает вопрос, связан ли высокий уровень КБГ-К в растениях удаленными ветвями с усиленным биосинтезом или, скорее, с уменьшением его ферментативной трансформации в другие каннабиноиды. Поскольку концентрации всех других каннабиноидов в этом методе были значительно снижены (на 25-40%) по сравнению с контролем, это означает, что увеличение КБГ-К при этой обработке является результатом ингибирования (подавления) метаболической активности по каннабиноидному пути.
Вопреки распространённому мнению производителей каннабиса, которые считают концентрацию каннабиноидов в первичном соцветии («колы») наивысшей в растении, мы сообщаем здесь, что для большинства обработок концентрации большинства каннабиноидов были выше в точках II и III, чем в апикальной меристеме точки I:
Точки отбора проб соцветий на растении
Поскольку на концентрацию каннабиноидов в соцветии влияют экзогенные (экологические) факторы, а также эндогенные эффекты развития / местоположения, разницу между методами обработки следует оценивать с учетом обоих этих аспектов. С точки зрения микроклимата, участки I, II и IV находятся в верхней части растения и сильно подвержены воздействию света и практически не затеняются другими растениями или ветвями. С другой стороны, местоположение III также сильно экспонируется, но может быть затенено соседними растениями в ряду или растениями из параллельных рядов в определённые часы фотопериода. Ожидается, что, как и в случае с интенсивностью света, в этих местах будет преобладать циркуляция воздуха, которая поддерживает одинаковую температуру и влажность в точках I и II, но практически не затрагивает точки III и IV. Следовательно, одни лишь изменения окружающей среды не могут объяснить выявленные вариации концентраций.
Несмотря на то, что во всех других точках отбора соцветий вариации интенсивности света практически не влияли на вторичный метаболизм, концентрации каннабиноидов в точке V были значительно ниже во все случаях примерно на 40%. Поскольку в этом месте уровни освещённости были очень низкими, свет, по-видимому, является ограничивающим фактором для вторичного метаболизма.
Чтобы оценить, влияют ли исследуемые обработки на степень изменчивости и имеют ли они потенциал для стандартизации химического потенциала, мы проанализировали влияние обработок на однородность химического профиля внутри растения.
Высокая степень однородности достигается за счёт химически похожих соцветий независимо от средней концентрации каждого каннабиноида при обработке. Анализ выявил несколько закономерностей:
Все обработки, кроме «двойной обрезки» и «BBLR», улучшили химическую однородность по сравнению с контрольной группой
Обработка BBLR дала наименее однородный результат
Удаление всех вторичных ветвей в течении вегетационного периода делало каннабис наиболее химически однородным
Оценка однородности профиля каннабиноидов
Чтобы сравнить реакцию отдельных каннабиноидов на методы изменения архитектуры, мы измерили средние концентрации каждого каннабиноида в растениях. Они обозначены в рисунке ниже:
Различия в производстве каннабиноидов в зависимости от архитектуры растения
Средние концентрации большинства идентифицированных каннабиноидов (КБД-К, ТГК-К, КБВ-К, ТГВ-К и КБХ) имели сходный, но не идентичный ответ на обработку архитектуры растения, в то время как реакция КБГ-К значительно различалась.
В целом, средние концентрации всех каннабиноидов, кроме КБГ-К, соответствовали тенденции Дефолиация = BBLR + Дефолиация > Двойная обрезка > Контроль = Одинарная обрезка > «BBLR» = Удаление вторичных ветвей > Удаление всех ветвей
Соцветия и урожай каннабиноидов
Накопление свежей биомассы различными органами растений в каннабисе представлено в рисунке ниже. Растения, прошедшие двойную обрезку, дали больше урожая, чем контрольные, а растения с удалёнными ветвями — меньше. Все другие методы обработки не оказали значительного влияния на биомассу урожая, демонстрируя пластичность развития репродуктивного роста в ответ на инвазивные изменения архитектуры побегов.
Биомасса соцветий, стеблей, веерных листьев и листьев соцветий каннабиса
Наивысший урожай каннабиноидов был достигнут в больше степени за счёт увеличения урожайности соцветий при двойной обрезке, чем за счёт увеличения концентрации каннабиноидов при дефолиации.
Выводы
Выращивание крупных растений позволяет производителям увеличить урожай каннабиса, но информация о свойствах и химической однородности такого урожая отсутствует. Поскольку влияние размера растений и архитектуры растений на микроклимат в побегах взаимосвязано, исследователи сосредоточили внимание на взаимосвязи между обработками архитектурных манипуляций и пространственной стандартизацией профиля каннабиноидов. Исследование было направлено на оптимизацию структурных манипуляций растений для повышения количества урожая и химического качества.
Результаты показали, что двукратная обрезка растений во время выращивания была оптимальной практикой для увеличения урожайности, а другие виды обработки снижали или не влияли на количество урожая. Хотя некоторые изменения химического профиля были вызваны обработками, и они в целом следовали следующей схеме:
Дефолиация = BBLR + Дефолиация > Двойная обрезка > Контроль = Одинарная обрезка > BBLR = Удаление вторичных ветвей > Удаление всех ветвей
Влияние этих химических изменений на общую продукцию каннабиноидов в растении было вторичным по отношению к эффектам биомассы урожая цветков. Следовательно, манипуляции с архитектурой могут использоваться для увеличения урожайности биомассы, но увеличения каннабиноидов следует достигать другими способами. Наблюдалось значительное уменьшение светопропускания вдоль побега, а степень пространственной стандартизации профиля каннабиноидов коррелировала с влиянием архитектуры растения на проникновение света в нижние части полога.
Это говорит о том, что низкая доступность света в нижней части растения является мощным фактором, способствующим сокращению пространственной химической однородности. Повышение урожайности, достигаемое при выращивании крупных растений, достигается за счет низкой химической однородности растения. Эти результаты играют важную роль в разработке оптимизированных протоколов выращивания для индустрии каннабиса, чтобы гарантировать пациентам высококачественный медицинский продукт.
Оригинальное исследование: mdpi.com
Подготовил: @Nimand
Еще почитать:
Связь NPK с урожайностью каннабиса при беспочвенном выращивании
Каннабиноиды и терпены: способы увеличения вторичных метаболитов каннабиса
Видео: Освещение для каннабиса: FAQ и мифы. Лекция на русском
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}