Поиск сообщества
Показаны результаты для тегов 'физиология растений'.
-
 Всего было написано двенадцать томов. Нам же интересны будут первые три, в которых описываются общие для всех растений процессы. Том первый: Физиология растительной клетки. Фотосинтез. Дыхание. Скачать в PDF с гугл-диска (493 стр.) Содержание тома в спойлере: Том второй: Минеральное питание. Рост и развитие. Эмбриогенез и ортогенез. Скачать в PDF с гугл-диска ( 481 стр.) Содержание в спойлере: Третий том: Физиология водообмена растений. Устойчивость растительных организмов. Природа иммунитета. Скачать в PDF с гугл диска (411 стр.) Содержание тома в спойлере: Примеры страниц: Читать всем любознательным. Действительно всеобъятный труд отвечающий практически на все вопросы по физиологии растений. Более детального труда на русском языке просто нет.
Всего было написано двенадцать томов. Нам же интересны будут первые три, в которых описываются общие для всех растений процессы. Том первый: Физиология растительной клетки. Фотосинтез. Дыхание. Скачать в PDF с гугл-диска (493 стр.) Содержание тома в спойлере: Том второй: Минеральное питание. Рост и развитие. Эмбриогенез и ортогенез. Скачать в PDF с гугл-диска ( 481 стр.) Содержание в спойлере: Третий том: Физиология водообмена растений. Устойчивость растительных организмов. Природа иммунитета. Скачать в PDF с гугл диска (411 стр.) Содержание тома в спойлере: Примеры страниц: Читать всем любознательным. Действительно всеобъятный труд отвечающий практически на все вопросы по физиологии растений. Более детального труда на русском языке просто нет. -

Агротехника, Физиология и оборванные листья
borroo опубликовал тема в Тренировки и гроверские методы
У каждого растения своя агротехника, связанная в основном с требованиями к конечной продукции и физиологией самого растения - какое оно: однолетнее, двухлетнее, многолетнее, длиннодневное, короткодневное, тепло - или влаголюбивое и т. д. Агротехника-система приемов возделывания культурных растений Например на кусте обычного синего баклажана оставляют четыре баклажана. Остальные завязи удаляют. Тогда баклажаны получаются не слишком большие и не слишком мелкие, как если бы оставили три или пять. Дыню в холодном климате оставляют одну и подвешивают в мешочке над землей. Она не должна касаться земли, чтобы в холоде не загнить и не должна оторваться от плети, когда вырастет. А если пожадничать, и оставить две завязи - вы не получите ни одной дыни, так как то, что вырастет, вы не будете есть, один корень две дыни не вывезет. Наше растение Оно однолетнее. Его можно долго держать на веге, но выращивают его как однолетнее. Оно не делает запасов на следующий год в виде кочерыжки или луковицы, т.е. переводя световой режим на цветение, вы включаете ему обратный отсчет. Оно умрёт, сенсимилья (в природе семена) его высосет и убьет в итоге. Самое слабое звено у нашего растения - это корни. Они всегда снабжаются по остаточному принципу. Им нужно расти весь цикл: сначала на корне появляются корневые волоски (только они могут всасывать ионы вместе с водой). Средний срок их жизни - три дня, потом большинство отмирает, и только очень некоторые покрываются лубом и становятся настоящими корнями (волоски должны продолжать расти всё время). Ионы, попавшие в корень, дальше двигаются по транспортной системе вверх. У растения транспортных систем две: одна сеть сосудов расположена под корой, это лыко или волокно конопли называется ксилема. Она состоит из трубочек, эти трубочки состоят из одеревенелых клеток, по ним ионы двигаются вверх вместе с восходящим потоком сока. Сок этот двигают, так называемые, концевые двигатели, верхний и нижний. Подробно про работу нижнего не буду, если растение срубить - выделится на срубе жидкость – пасока. Это работа нижнего концевого двигателя, он слабее верхнего. Верхний концевой двигатель работает за счёт испарения воды через устьица - транспирации и выделения воды из желёзок по краю листа-гуттации. Второй пучок волокон расположен внутри стебля, называется флоэма. По ней вещества двигаются сверху к корням. Она состоит из живых клеток, по ней сок не двигается, по ней перемещаются вещества с затратой энергии. Вообще ксилема считается проводником ионов, а флоэма органических веществ, но по ксилеме могут двигаться аминокислоты и по флоэме калий, фосфор, а так же между ними есть сообщение - некоторые вещества могут перемещаться радиально, из флоэмы в ксилему и наоборот. Это явление называется транслокацией (это выгодно некоторым веществам - не надо делать круг по растению). Ионы из корня попадают по ксилеме в листья и меристемы надземной части. Меристемы - это область усиленного деления клеток, где происходит основной рост массы. На веге - это точки роста и молодые листья, на цветении - это шишки (там больше всего гормонов, клетки быстрее всего делятся, там больше всего расходуется продукция фотосинтеза). И только во вторую очередь снабжаются по флоэме питанием меристемы корня, при этом еще энергию надо тратить, то есть, опять же продукцию фотосинтеза. Продуктивность фотосинтеза Это самое главное. Здоровая корневая система + правильное питание удобрениями, толстая у основания кочерыжка (хорошо развитая транспортная система) , правильный климат + хорошее освещение + дополнительная подача СО2 – всё это делается, чтобы превратить углекислый газ и воду с помощью света в одно вкусное, но не полезное для человека в чистом виде вещество. Чем больше в граммах сахарозы вы получите за цикл, тем на большее количество шишек вправе рассчитывать. Из сахарозы в растении делаются все остальные органические и пластические вещества, из которых лепятся новые ткани растения и берётся энергия для энергозатратных процессов. Зачем можно отрывать листья? 1. Сформировать толстый ствол, остановить рост вверх. Но есть несколько менее варварских способов это сделать, можно прочитать например здесь (знание английского обязательно) 2. Смоделировать нападение насекомых или животного, заставить растение выработать иммунитет и выделить еще смолы. Но есть препараты с действием более сильным, безвредные для человека и оставляющие листья в покое, на основе хитозана. Bud Factor X, например. Почему нельзя отрывать листья? Что происходит, когда вы обрываете листья? Растение выделяет гормоны старения и стресса - этилен и абсцизовую кислоту. Растение под действием абсцизовой кислоты закрывает устьица и прекращает транспирацию и фотосинтез. Если сильно оборвали – на несколько дней свет будет пропадать зря. Растение под действием этилена будет наращивать ствол и перераспределять токи в изменившемся объеме. Если оборвёте очень много - этилен состарит нижние листья, а абсцизовая кислота заставит их отвалиться. Растение ре-утилизирует органические вещества из этих листьев, фотосинтеза-то нет на попытку отрастить новые листья. А так как основную массу органических веществ отнимает меристема, в условиях дефицита оных у вас рвётся самое слабое звено – корни (им ничего не достается). Корневые волоски надолго перестают расти. Дальше вы ломаете верхний концевой двигатель, при том, что нижний двигатель вы тоже уже сломали, гуттация и транспирация в прежнем объеме восстановится, когда новые листья отрастут (если отрастут). Дальше. Все листья на растении разные: верхние с толстым защитным слоем из воска-кутикулы настроены на сильный свет и предохраняют другие, те, что в глубине куста от ожогов, их хлоропласты настроены собирать слабый рассеянный свет (прямой их обжигает). Дальше . Фоточувствительность. Растение эволюционировало на одном месте, куда бы семечко не попало - в кусты или другую тень, у него есть механизм, как расположить листовую мозаику так, чтобы из любой ситуации выжать максимум света, поворотный механизм, фототропизм. Самые фоточувствительные лопухи у ствола. Очень чувствительный механизм – наш глаз, еле различает день-ночь по сравнению с ним. Если вы считаете, что куст что-то неправильно освещает, то нет. Дальше. Вы лишаетесь сахара. Не только того, который лист производит, но и того, который растение могло бы взять на ре-утилизацию. Для повторного использования могут использоваться и некоторые ионы, т.н. подвижные, чей недостаток проявляется на старых листьях. Ими растение жертвует, чтобы восполнить недостаток - молибден, магний, азот и т.д. Их запас вы тоже теряете. Дальше. Затрудняется транслокация. Она происходит в листьях, где флоэма и ксилема расположены ближе всего - в жилке листа. Дальше вы просто можете заразить растение через множество открытых ран. Заключение Наше растение по агротехнике относится к растениям, у которых удаляются лишние точки цветения. Если на предцвете в низу или внутри куста из пазухи листа лезет зародыш шишки на длинной палочке - их стоит удалить, из них путного ничего не будет, даже если дать близлежащим листьям свет (цветам свет вообще не нужен). Формировать куст с как можно более короткими расстояниями между нодами (узлами), по удочкам тяжело вещества гонять, отсекать затененные ветви, они потом начнут атрофироваться, если совсем в тень попадут. Сразу замазывать раны антисептиком для ран на растениях - Садовым Варом или РанНет. Обсудить на форуме- 79 ответов
-
- 68
-

-
- физиология растений
- агротехника
- (и ещё 1 )
-
Реакция на фотопериод -адаптация растений к сезонным изменениям среды,если растение обитает в каких-то меняющихся климатических условиях,в умеренных и полярных широтах например сокращение длинны дня сигнализирует наступление холодов,в местах потеплее может означать приближение влажного сезона.Это приспособление к конкретным условиям,в которых эволюционировало растение, сохраняется при перенесении его выращивания в другой климат. Фотопериодом регулируется: индукция цветения, формирование запасающих органов-клубней,луковиц, сезонный переход в состояние покоя для многолетних растений вегетативное размножение- усы у земляники,расселительные побеги Наиболее изучена фотозависимая индукция цветения длиннодневные зацветают,когда длинна дня превышает какой-то критический период,короткодневные-наоборот.Короткодлиннодневным и длиннокороткодневныемдневным мало только индуктивного периода,им нужно чередование периодов,это первоцветы и растения,цветущие осенью.Фотонейтральные-растения,зацветающие за счет других факторов,возраста или достаточной биомассы например-это пять больших групп и две маленькие.Среднедневные цветут на среднем дне,это некоторые кактусы,амфифотопериодические на определенной длине дня не цветут,на всякой другой длине дня цветут. Также реакция может быть количественной и качественной,например арабидопсис,модельное растение физиологов растений,цветет как на коротком,так и на длинном дне,но на длинном дне цветет интенсивнее,это реакция количественная.У фотопериодических сортов конопли реакция качественная,на длинном дне цветение прекращается. Явление фотопериодизма открыли в 20-ые годы в США на табаке сорта Мерилэнд,произошла случайная мутация и табак на длинном дне не цвел и вырастал очень большой.А при перенесении зимой в теплицу на короткий день зацветал.Если ему прерывали ночь,цветение прекращалось,параллельно оказалось существует другая группа растений с симметричными реакциями. В СССР темой индукции цветения занимался Михаил Христофорович Чайлахян с 30-ых годов прошлого века и до своей смерти в 91 г,многие его опыты стали классическими,а его гормональная теория цветения растений получила повсеместное признание. Чайлахян решил выяснить,какой орган воспринимает фотопериод.Свету открыты стебли,листья и меристемы побегов(точки роста) и он поставил простой эксперимент,удалил все молодые листья у короткодневных растений хризантема индийская,одному закрыл светонепроницаемым мешком верхушку побега,другому старые листья,плюс пара контрольных растений,полностью открытое и полностью закрытое Дело происходило летом на длинном дне,Чайлахян приходил на работу и на 8 часов снимал мешки,имитируя короткий день.Логика эксперимента проста,если цветение инициируется меристемой или листьями,это будет понятно,если стебель,не произойдет ничего.И вуаля Флоральный сигнал формировался в старых "компетентных" листьях,а после передавался в меристему стебля.Причем сигнал передавался в том числе замысловатым путем,в одном из экспериментов Чайлахян разрезал скальпелем растение надвое,оставлял общим только кончик корня.В одной половинке в листе формировался флоральный сигнал,передавался вниз к корню,попадал в кончик,поднимался вверх и вызывал во второй половинке цветение.Часто он использовал для опытов Периллу это живучее растение,его удобно резать,производить трансплантации.Прививая индуцированные листья топинамбура(короткодневное растение) к подсолнечнику(нейтральное) он сумел вызвать его цветение.Таким образом он выяснил,что предполагаемое вещество,он назвал его флориген,может перемещаться вверх и вниз по растению и видонеспицифично,т.е. вызывает цветение у разных видов растений. Когда Чайлахян начинал опыты,был открыт один гормон,ауксин.В 50-ые были открыты гиббериллины,Чайлахян долго их рассматривал компонентами флоригена,но они вызывали цветение только длиннодневных растений.В добавок к уже существующим теориям цветения(азотно-углеводной теории Клебса например) добавились мелантоиновая,синтетическая и другие,гормональная была одной из них.Каждая что-то объясняла,но флориген оставался неуловим до 21-ого века. И вот наконец в 2005-ом году были открыты не гормональные вещества,а белки со свойствами гормонов массой 23 кДа.Это оказался транскрипционный фактор,белок связанный с участком ДНК.Участок этот называется флоральный локус Т(flowering locus T),FLT или FT,а белок назвали в честь Чайлахяна флоригеном. Этот участок ДНК(FLT) можно переносить с помощью генной инженерии.Для этого используется транспорт,вирус капустной мозаики,в которую вместо генов,кодирующих белки оболочки,вставлены гены интереса,в данном случае производящие флориген.Или использовать устройство,которое называется генная пушка.Она скорее не пушка,а пистолет,первая генная пушка из строительного пистолета была сделана. На пулю наносятся частички металла с ДНК,происходит выстрел или сейчас это сжатый газ,пуля натыкается на перегородку с отверстием,останавливается,а частички металла застревают в растении. В 2011-ом году таким образом удалось вызвать цветение у саженцев яблони первого года речь конечно о получении яблок не идет,но можно собрать пыльцу.Раньше чтобы быть успешным селекционером плодовых деревьев,мало было хорошо разбираться в селекции,нужно было прожить довольно длинную жизнь.Современные сорта яблок впервые цветут через 6-8 лет,Мичурин первых цветов яблонь ждал 10 лет http://elementy.ru/n...tskimi_uchenymi https://www.nkj.ru/news/20648/ http://www.rusplant....eke6bvoqp3cq363
-
В данной статье я хочу рассказать о том, как растение потребляет питательные вещества на самом базовом уровне - а именно о том, как питательные вещества поступают в клетку растений... Хочу сразу предупредить особо рьяных критиков: в данном теме я не рассматриваю эндо- и экзоцитоз, то есть регуляцию растениями цельных незаряженных молекул питательных веществ. В какой-то мере по этой причине информация здесь может считаться неполной, но тем не менее, я считаю, что данный текст будет полезен тем, кто хочет прикоснуться к настоящей научной агрономии. Мембранный транспорт Мембранный транспорт – это общее название всех процессов транспортировки веществ в клетку и из клетки через клеточную мембрану. Мембраны обладают избирательной проницаемостью, что означает, что только определенные молекулы и частицы способны проникать через нее. Клеточная мембрана устроена намного сложнее, чем любая искусственная мембрана, в первую очередь потому, что мембранный транспорт клетки обеспечивают белки переносчики, которые изменяют проницаемость мембраны в сторону большей или меньшей проницаемости для определенных растворов. К сожалению, это не означает, что растение может самостоятельно регулировать проницаемость клеточной мембраны – только то, что проницаемость мембраны может быть разная даже в похожих клетках. Мембранный транспорт может осуществляться по или против градиента электрохимического потенциала. Электрохимический потенциал можно представить, как давление на клеточную стенку либо изнутри, либо снаружи, но не физическое давление, как давление воды на плотину, а как притяжение заряженных частиц к одной из сторон. Движение к более низкому электрохимическому потенциалу не требует затрат энергии, и называется пассивным транспортом. Движение против градиента требует затрат энергии и называется активным транспортом. Здесь очень важный момент, следует отличать градиент электрохимического потенциала от градиента концентрации (водного потенциала). В сообщающихся растворах все элементы стремятся к равной концентрации независимо от других. Градиент концентрации указывает на стремлении к равной концентрации растворителя, то есть воды. Когда мы измеряем электропроводимость раствора, мы измеряем концентрацию ионов в нем, но это не говорит о том, больше ли, например, ионов калия в одном растворе, чем в другом. Но это говорит о том где больше концентрация воды, и в какую сторону направлен ее потенциал. То есть, пользуясь кондуктометром мы на самом деле измеряем концентрацию воды. Но если в первом сосуде концентрация ионов, например, кальция будет выше, чем во втором, то ионы будут перемещаться из первого в сторону второго, независимо от градиента концентрации между растворами Пассивный транспорт Прежде чем мы рассмотрим пассивный транспорт, поговорим о том, как вообще устроена клеточная мембрана или, как она еще называется, плазмалитическая мембрана. Она представляет собой двойной слой молекул фосфолипидов, состоящих из гидрофильной (водопроницаемой) головки и гидрофобного (водонепроницаемого) хвоста. Молекулы расположены головками наружу – в клетку и окружающую ее среду, а хвосты образуют гидрофобный слой, ограждающий клетку от диффузии между клеточной жидкостью и раствором снаружи. Плотный билипидный слой прерывается белками – каналами и переносчиками, которые и обеспечивают основный обмен элементами между клеткой и окружающей средой. Осмос Для начала следует понять, как именно вода поступает в клетку растения. Вода всегда перемещается по градиенту концентрации – в тот раствор, где ее концентрация меньше. В нормальных условиях концентрация воды выше внутри клетки, чем снаружи, но как вода преодолевает гидрофобный слой плазматической мембраны? Помогают ей в этом специальные белки – аквапорины. Мономеры аквапоринов образуют водяную пору, сужающуюся структуру, через которую вода проходит как песок через песочные часы. Аквапорины не пропускают заряженные частицы – ионы питательных веществ, однако помимо воды через них могут также проходить небольшие незаряженные частицы - мочевина, глицерол, борная и кремниевая кислоты, перекись водорода, а также газы (CO2 , NH3 , NO). Читайте также: Детка, ты просто осмос! Диффузия Через мембрану клетки могут беспрепятственно проходить только очень маленькие неполярные молекулы, такие как газы O2 и CO2. Билипидный барьер надежно защищает клетку от неконтролируемого проникновения любой жидкости или питательных веществ. Тем не менее, диффузия может произойти вследствие повреждения клеточной мембраны. Но это скорее аномалия, поэтому мы ее рассматривать не будем, так как ее влияние на питание растения ничтожно. Облегчённая диффузия Облегчённая диффузия (название то какое) – это диффузия через специальные каналы, которые клетка заботливо и заблаговременно оставила в клеточной мембране. Это не просто дырки в мембране, это совершенно другой процесс. В его основе лежит унипорт. Небольшое отступление – унипорт, симпорт и антипорт – так можно разделить мембранные транспортеры по количеству проходящих через них ионов и их направлению. Унипорт – это канал, в который проходит один ион в одном направлении. Симпорт – два иона в одном и том же направлении. Антипорт – два иона одновременно проходят через канал в противоположных направлениях. Так вот, белки, через которые проходит облегченная диффузия – это унипорты. В отличии от обычной диффузии транспорт через каналы отличается избирательностью – через один вид канала может проходить только лишь определенный вид ионов. Также отличия есть и в зависимости скорости транспорта от разницы концентраций и возможности подавления работы каналов специальными ингибиторами. Активный транспорт Описывая унипорт, мы говорили об ионных каналах, но ионные каналы не единственные мембранные транспортеры в клетках. Помимо каналов существуют еще и белки-переносчики. Белки переносчики захватывают ион питательного вещества с одной стороны мембраны, а затем меняют форму и «выталкивают» ион с другой. Ну а ионные каналы это эмм… каналы. Они могут открываться и закрываться и ионы спокойно проходят через них, когда они открыты. Разница не настолько мала, как может показаться: белки-переносчики быстро достигают максимума скорости транспорта при достижении определенной концентрации – они физически не могут транспортировать больше ионов, в то время как зависимость транспорта через каналы от концентрации более ярко выражена. Белки-переносчики дают больший контроль над протеканием транспорта, а ионные каналы – большую скорость: проницаемость канала на три порядка выше, чем белка-переносчика и на 11 порядков выше, чем простая диффузия через мембрану. Наконец, мы переходим к активному транспорту. Как мы помним, активный транспорт – это перемещение ионов против электрохимического градиента, то есть туда, где их уже больше. Поверхностное представление о физике дает нам понимание, что такие вещи не могут происходить бесплатно. Так и есть – на осуществление активного транспорта требуется энергия, которую растение получает из распада молекулы АТФ – аденозин-три-фосфата. Активный транспорт делится на первично-активный и вторично-активный. Разницу обсудим позже, а пока поговорим о первично-активном транспорте В основе первично-активного транспорта лежат ионные помпы. Ионные помпы – это белки, которые связывая различные ионы, способны выталкивать их в разных направлениях независимо от градиента электрохимического потенциала. По сути это насосы для катионов. Их еще называют АТФ-азами, по запускающей их работу молекуле. Давайте подробнее рассмотрим, как работают ионные помпы на примере протонной помпы. Ион водорода H+ (гидроксоний) внутри клетки закрепляется на внутренней стороне помпы. Молекула АТФ прикрепляется к рецепторам на помпе внутри клетки и распадается. Высвобожденная энергия изменяет форму белка таким образом, что гидроксоний оказывается снаружи клетки. Как только ион водорода отсоединяется от помпы, она возвращается в изначальное состояние. Вы спросите, зачем вообще клетка выпускает ионы водорода в окружающую среду? А дело в том, что это – основной механизм получения питательных веществ растениями. Выпуская кучу ионов вплотную к клеточной стенке, растение уменьшает концентрацию других ионов возле нее, то есть как бы разбавляет их своими ионами водорода. Это создает градиент электрохимического потенциала по направлению к клетке, где потом они захватываются средствами вторичного активного транспорта, но об этом чуть ниже, а пока о помпах. Кальциевые помпы (или Ca2+-АТФазы). Клетка может выпускать ионы кальция в окружающий раствор помпами, а потом забирать их через каналы. Существует целая система активного транспорта ионов кальция, и отнюдь не только потому, что растение в нем очень нуждается как в строительном материале. Кальций – основной элемент вторичной сигнальной системы растения. Про это можно отдельную статью написать, но сейчас я расскажу вкратце. Клетки способны способны улавлить мельчайшие изменения концентрации ионов кальция, а также кальций легко взаимодействует с белками. Это связано с электрохимическими свойствами кальция. Кальциевые помпы закачивают и выкачивают кальций из окружающей среды и складов кальция внутри клетки (которые называются эндоплазматические ретикулумы, но можете развидеть это), удерживая низкий, но контролируемый уровень иона в цитоплазме, тем самым контролируя скорость протекания белковых реакций. Тяжелые металлы (такие как Fe, Cu, Mn, Zn и Mo) транспортируются в клетку большой группой транспортеров, известной как P1B. Так как все эти металлы могут быть токсичны в больших количествах, растение активно регулирует их содержание в клеточной жидкости, выталкивая излишки наружу. Все перечисленные выше АТФ-азы относятся к АТФ-азам P-типа (P здесь значит фосфор – по названию активирующего их работу элемента). Помимо них есть АТФ-азы V-типа и F-типа, первые используются для переноса веществ внутри клетки, а вторые для производства фотосинтеза. Но нам они не интересны, по крайней мере в рамках этой статьи. Отдельной группой стоят так называемые ABC-переносчики. Это огромное семейство транспортеров, которые обеспечивают мембранный транспорт пептидов, аминокислот, сахаров, липидов, ионов металлов, полисахаридов, вторичных соединений, неорганических кислот и других молекул. ABC-белки могут транспортировать фенольные соединения и алкалоиды, терпеноиды и воска, стероиды и продукты распада хлорофиллов, гербициды и тяжелые металлы. АВС-переносчики участвуют в полярном транспорте ауксина, катаболизме липидов, устойчивости к болезням, работе устьиц и детоксикации. Короче, это один из главных регуляторных механизмов растения, особенно тех, что касаются сложных органических и неорганических молекул. Тем не менее эти белки не имеют особо важной роли в питании растения, их функции другие. Отличительной особенностью их от переносчиков P-типа является наличие двух дополнительных частей (ABC-доменов) внутри клетки, что делает механизм их работы слегка другим. В открытом состоянии переносчик связывает специфичную для него молекулу, затем он закрывается, и к ABC-доменам присоединяются две молекулы АТФ, что приводит к видоизменению белка и выбросу связанной молекулы наружу. Вторично-активный транспорт Теперь, когда самое сложное уже позади и вы, я надеюсь, прочитали и даже поняли предыдущий текст, я расскажу, зачем растению нужна протонная помпа. Дело в том, что выкачать из клетки ион водорода оказывается легче и менее энергозатратно, чем транспортировать более тяжелые элементы внутрь. Поэтому клетка, выбрасывая катион водорода в околоклеточный раствор, создает электрохимический градиент этого катиона по направлению внутрь клетки. Я вкратце говорил, что такое электрохимический градиент вначале статьи, сейчас хочу рассказать подробнее. Электрохимический градиент – это по сути сумма электрического и химического градиента. Он выражается в количестве энергии, которую нужно приложить, чтобы передвинуть ион против градиента или количество энергии, которое высвобождается при движении иона по градиенту. Согласно закону энтропии, энергия стремится уравновесить себя в пространстве. Чем больше концентрация веществ с одной или другой части сообщающихся сосудов, тем больше их суммарная энергия. Поэтому ионы всегда двигаются в сторону меньшей концентрации. Это химический градиент. С другой стороны, заряженные частицы создают электрическое поле. Если в одном растворе положительно заряженных частиц больше чем в другом, то они будут перемещаться вдоль создаваемого ими магнитного поля до тех пор, пока заряды не уравновесят друг друга. Это электрический градиент. Электрохимический градиент – это сумма химического и электрического градиента. Так вот, вторично-активный транспорт для протонных помп выполняют белки-переносчики, которые являются симпортами (или котранспортерами). Помните, ранее я рассказывал, что мембранные транспортеры делятся на унипорты, симпорты и антипорты? Так вот, симпорт двигает два иона внутрь клетки, используя электрохимический градиент одного из них. Вот как это происходит: протонная помпа создает электрохимический градиент, активно выкачивая катионы водорода из клетки. Катион питательного вещества присоединяется к белку переносчику. Затем, к нему же присоединяется катион водорода. Энергия, высвобождаемая прохождением катиона водорода внутрь клетки, используется для продвижения внутрь клетки иона питательного вещества против его градиента концентрации. Как видите, энергия на активный транспорт питательного вещества затрачивается не напрямую, а опосредованно протонной помпой. Другой тип белков вторичного активного транспорты – это антипорты (или обменники). Суть та же, что и в симпортах, только в антипортах молекулы транспортируются в разных направлениях. В связке с протонной помпой и электрохимическим градиентом H+ антипорты чаще служат для выталкивания элементов наружу, а не внутрь клетки. Учитывая важность протонный помпы для жизнедеятельности растения, вы должны понимать важность pH раствора, который как я надеюсь вы знаете, является мерой концентрация катиона водорода H+. При слишком высоком pH, то есть малой концентрации катионов водорода, энергии электрохимического градиента может не хватить для переноса вещества в клетку средствами вторично-активного транспорта, а при слишком низком pH, то есть высокой концентрации Н+, катионы водорода могут забивать трансмембранны поры. Мембранный транспорт большинства катионов у растений осуществляется за счет энергии электрохимического градиента ионов H+. Таким путем в растительную клетку или из нее могут транспортироваться такие катионы, как K+, NH4+ Na+, Ca2+ и Mg2+. Белки-переносчики калия – это три группы, два симпорта и один антипорт (HKT, KT/KUP/HAK и CPA соответственно, если захотите узнать о них поболее). Антипорт калия менее специфичен, чем симпорты, через него могут выталкиваться и другие протоны, но это то – что растению и нужно – выталкивать излишки. Аммоний транспортируется переносчиками симпортами класса AMT. Их активность пропорциональна электрохимическому градиенту Н+. Помимо первично-активных кальциевых помп, растение избавляется от излишков кальция в цитоплазме с помощью Н+/Са2+ -обменников. Натрий, который не слишком нужен растению, транспортируется похожим H+/Na+ -антипортом. Транспорт магния осуществляется магниевыми помпами и обменниками, и в целом похож на транспорт кальция. Большая часть информации о типах мембранных транспортеров появилась из расшифровки генома растений и принадлежит последним двум десятилетиям. Я хочу сказать, что вероятнее всего мы еще не знаем о существовании многих видов транспортных каналов. Транспорт аниов протекает в основном симпортом с протоном водорода. Таким путем в растительную клетку поступают фосфат, нитрат и сульфат. Переносчик фосфора в плазматической мембране – это белки семейства Pht1. Pht1. Нитраты переносят NRT1 и NRT2 –симпортеры, сильно различающиеся скоростью и объемом транспорта. Сульфатные переносчики состоят из 3 типов, два из которых отвечают за переносом вещества в корневой, а третий – через лист во флоэму. Ионные каналы Вернемся к нашим каналам. Вкратце я о них уже рассказывал, теперь поподробней. Как вы уже, наверное, поняли, ионные каналы — это основной механизм пассивного транспорта в клетке – облегчённой диффузии. Избирательность канала обусловлена формой его селективного фильтра – через один вид канала может проходить один (хотя иногда и несколько) вид иона. Но каналы еще должны уметь открываться и закрываться, независимо от присутствия нужного иона, иначе их поток будет неуправляем. Положение открытия/закрытия канала (воротный механизм) регулируется набором факторов, зачастую несколькими из них. Белок может открыться из-за присоединившейся к сенсору канала молекулы (лиганда). Лиганд, присоединяясь к каналу, меняет его форму, и он открывается. Канал также может открываться из-за величины мембранного потенциала, то есть электрическому напряжению мембраны, из-за pH, растяжения мембраны и света, последнее относится к клеткам надземной части. Выход иона из канала облегчается при появлении на входе другого иона из-за их электростатического отталкивания. Однако при высоких концентрациях может происходить насыщение канала из-за заполнения ионами его входа и выхода и, как следствие, его блокировка. Через селективные каналы мембраны могут проходить в обоих направлениях ионы К+, Са2+, в меньшей степени NH4+, Ba2+, Mg2+. Но если все бы было так просто мы бы никогда не смогли бы получить в растениях токсичности по определенным элементам. Помимо селективных в клеточной мембране растений также имеются неселективные каналы. Они способны пропускать Na+, NH4+, Mg2+, Zn2+, Li+, Cs+, Ba2+ и другие катионы. Анионные каналы работают также, как и катионные, с небольшим отличием в том, что на вход им нужно преодолевать электрический потенциал, созданный протонный помпой. Поэтому анионные каналы работают только как резервные средства при сильном дефиците анионов в растении. Так их основная функция – избавление от анионов при их переизбытке в цитоплазме. Но анионные каналы существуют не только для минеральных питательных веществ. Они могут выпускать малат и цитрат, продукты метаболизма растения, которые, являясь слабыми кислотами, контролируют уровень pH вокруг корня. Еще они выпускают органические соединения, которые связывают Al3+, токсичный для многих процессов растения. Выводы На основе этой информации вы можете сделать свои выводы. Вот вам мои. Растение обладает огромными возможностями в контроле поступаемых в него питательных веществ. Это – результат эволюции в миллиарды лет. Любое вещество, попавшее в клетку растения, может быть вытолкнуто наружу. Что это значит? Что не существует никакой идеальной формулы питательного раствора, существует оптимальный диапазон для каждого питательного вещества. Идеальный раствор – это тот, в котором растение затрачивает минимум энергии на получение питательных веществ и минимум энергии на то, чтобы избавляться от питательных веществ, выбрасывая их в околоклеточный раствор. Можем ли мы высчитать объем необходимого питания используя информацию о мембранных каналах? К сожалению, нет. Во-первых, механизм работы многих транспортеров до сих пор недостаточно понятен. Во-вторых, точную информацию о количестве и типах, присутствующих в растении каналов, мы можем собрать, только лишь расшифровав геном растения. Геном каннабиса не расшифрован. Большая часть информации в этой статье построена на физиологии совсем другого растения, этакой лабораторной мыши для ботаников – резуховидки Таля, также известной как арабидопсис. Но кое-что мы все же определить можем. pH среды важен настолько, насколько он важен в прикорневой области, ризосфере. Растение может легко регулировать pH прикорневой в субстратах типа земли или кокоса. В гидропонных системах типа ДВЦ, в которых растение продолжительное время находится в контакте с раствором, регуляция pH в основном ложится в руки гровера. Общая концентрация питательных веществ важна не меньше, чем их соотношение. При высокой концентрации растению становится труднее пользоваться ионной помпой, так как оно удерживает мембранный потенциал на определенном уровне. С другой стороны, высокая концентрация благоприятно сказывается на пассивном притоке питательных веществ в растение, хотя и ухудшает работу каналов «на выход». Слишком низкая концентрация питательных веществ может сказаться еще хуже, чем их избыток – растение останавливает рост. По моему мнению легкий передоз лучше, чем легкий дефицит. Но самый лучший вариант – держать концентрацию оптимальной. Что можно сказать о соотношении элементов? Я бы разделил все питательные элементы на две группы – те, которые растение привыкло видеть в избытке и те, которые оно привыкло видеть в дефиците. К первым я бы отнес калий, кальций, магний и серу. Это те элементы, которые находятся в избытке в большинстве почв. Калий – это единственный элемент растения, который может находиться в равновесной концентрации в клетке и окружающем растворе. Растению нужно много калия, его избыток может привести к проблемам электрохимического потенциала мембраны, но сам он совершенно не токсичен для растения. Растение обладает развитой системой регулировки содержания кальция в цитоплазме. Разница концентраций кальция внутри клетки и снаружи может составлять несколько порядков. Магниевые транспортеры во многом схожи с кальциевыми, а сера – это анион, и растение может легко избавиться от нее открыв анионные каналы и выбросив в ее в слегка положительно заряженную окружающую среду. Первая группа элементов это та, на которую я бы советовал обращать меньше внимания. Растение обладает механизмами, позволяющими регулировать их абсорбацию, главное, чтобы их было достаточно. Другое дело вторая группа, к которой я отношу азот и фосфор. Азот медленно восстанавливается в почве, он базовый элемент растения в накоплении биомассы и растение привыкло забирать его полностью. Несколько видов азота в растворе позволяет задействовать больше каналов и улучшить его абсорбацию и регуляцию. С фосфором проще, так как он анион, растению легче от него избавиться. Но я считаю, что растение не могло эволюционно развить высокоэффективные механизмы регулирования его содержания в клетке. Что я хочу сказать, так это то, что соотношение питательных элементов в растворе можно свести в большей степени к количеству азота, в меньшей – фосфора и в совсем небольшой к количеству других макроэлементов. Что касается микроэлементов, в большой концентрации они токсичны для растения, поэтому оно обладает механизмами удержания их в клетке в необходимой концентрации, даже когда разница концентрации с околоклеточным раствором составляет несколько порядков. Я бы советовал обращать внимание только на бор, который может поступать через аквапорины, хлор и натрий, присутствие которых в субстратах может быть чрезмерным для компенсаторных механизмов растения. Лучшей инструкцией как правильно кормить является ваш опыт и интуиция, которые вы должны «прокачать», прочитав эту статью. Следите за азотом, pH, и общей концентрацией. Спасибо всем, кто дочитал эту простыню до конца. Удачных гровов и больших шишек! Аминь! Статья победитель апрельского конкурса "Автор, жги" Читай также: Гроупедия - огромная энциклопедия по выращиванию. Постоянно пополняется интересными материалами. Гид по поливу растений Анионы и катионы: сбалансирована ли ваша система?
- 12 ответов
-
- 15
-
-
- физиология растений
- мембрана
- (и ещё 3 )
-
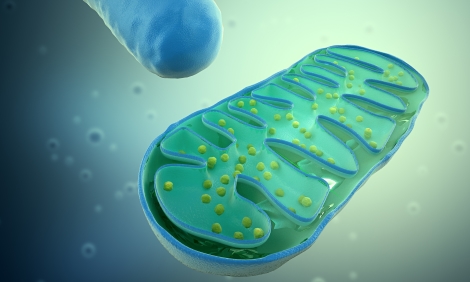 Хочу сразу предупредить особо рьяных критиков: в данном теме я не рассматриваю эндо- и экзоцитоз, то есть регуляцию растениями цельных незаряженных молекул питательных веществ. В какой-то мере по этой причине информация здесь может считаться неполной, но тем не менее, я считаю, что данный текст будет полезен тем, кто хочет прикоснуться к настоящей научной агрономии. Мембранный транспорт Мембранный транспорт – это общее название всех процессов транспортировки веществ в клетку и из клетки через клеточную мембрану. Мембраны обладают избирательной проницаемостью, что означает, что только определенные молекулы и частицы способны проникать через нее. Клеточная мембрана устроена намного сложнее, чем любая искусственная мембрана, в первую очередь потому, что мембранный транспорт клетки обеспечивают белки переносчики, которые изменяют проницаемость мембраны в сторону большей или меньшей проницаемости для определенных растворов. К сожалению, это не означает, что растение может самостоятельно регулировать проницаемость клеточной мембраны – только то, что проницаемость мембраны может быть разная даже в похожих клетках. Мембранный транспорт может осуществляться по или против градиента электрохимического потенциала. Электрохимический потенциал можно представить, как давление на клеточную стенку либо изнутри, либо снаружи, но не физическое давление, как давление воды на плотину, а как притяжение заряженных частиц к одной из сторон. Движение к более низкому электрохимическому потенциалу не требует затрат энергии, и называется пассивным транспортом. Движение против градиента требует затрат энергии и называется активным транспортом. Здесь очень важный момент, следует отличать градиент электрохимического потенциала от градиента концентрации (водного потенциала). В сообщающихся растворах все элементы стремятся к равной концентрации независимо от других. Градиент концентрации указывает на стремлении к равной концентрации растворителя, то есть воды. Когда мы измеряем электропроводимость раствора, мы измеряем концентрацию ионов в нем, но это не говорит о том, больше ли, например, ионов калия в одном растворе, чем в другом. Но это говорит о том где больше концентрация воды, и в какую сторону направлен ее потенциал. То есть, пользуясь кондуктометром мы на самом деле измеряем концентрацию воды. Но если в первом сосуде концентрация ионов, например, кальция будет выше, чем во втором, то ионы будут перемещаться из первого в сторону второго, независимо от градиента концентрации между растворами Пассивный транспорт Прежде чем мы рассмотрим пассивный транспорт, поговорим о том, как вообще устроена клеточная мембрана или, как она еще называется, плазмалитическая мембрана. Она представляет собой двойной слой молекул фосфолипидов, состоящих из гидрофильной (водопроницаемой) головки и гидрофобного (водонепроницаемого) хвоста. Молекулы расположены головками наружу – в клетку и окружающую ее среду, а хвосты образуют гидрофобный слой, ограждающий клетку от диффузии между клеточной жидкостью и раствором снаружи. Плотный билипидный слой прерывается белками – каналами и переносчиками, которые и обеспечивают основный обмен элементами между клеткой и окружающей средой. Осмос Для начала следует понять, как именно вода поступает в клетку растения. Вода всегда перемещается по градиенту концентрации – в тот раствор, где ее концентрация меньше. В нормальных условиях концентрация воды выше внутри клетки, чем снаружи, но как вода преодолевает гидрофобный слой плазматической мембраны? Помогают ей в этом специальные белки – аквапорины. Мономеры аквапоринов образуют водяную пору, сужающуюся структуру, через которую вода проходит как песок через песочные часы. Аквапорины не пропускают заряженные частицы – ионы питательных веществ, однако помимо воды через них могут также проходить небольшие незаряженные частицы - мочевина, глицерол, борная и кремниевая кислоты, перекись водорода, а также газы (CO2 , NH3 , NO). Читайте также: Детка, ты просто осмос! Диффузия Через мембрану клетки могут беспрепятственно проходить только очень маленькие неполярные молекулы, такие как газы O2 и CO2. Билипидный барьер надежно защищает клетку от неконтролируемого проникновения любой жидкости или питательных веществ. Тем не менее, диффузия может произойти вследствие повреждения клеточной мембраны. Но это скорее аномалия, поэтому мы ее рассматривать не будем, так как ее влияние на питание растения ничтожно. Облегчённая диффузия Облегчённая диффузия (название то какое) – это диффузия через специальные каналы, которые клетка заботливо и заблаговременно оставила в клеточной мембране. Это не просто дырки в мембране, это совершенно другой процесс. В его основе лежит унипорт. Небольшое отступление – унипорт, симпорт и антипорт – так можно разделить мембранные транспортеры по количеству проходящих через них ионов и их направлению. Унипорт – это канал, в который проходит один ион в одном направлении. Симпорт – два иона в одном и том же направлении. Антипорт – два иона одновременно проходят через канал в противоположных направлениях. Так вот, белки, через которые проходит облегченная диффузия – это унипорты. В отличии от обычной диффузии транспорт через каналы отличается избирательностью – через один вид канала может проходить только лишь определенный вид ионов. Также отличия есть и в зависимости скорости транспорта от разницы концентраций и возможности подавления работы каналов специальными ингибиторами. Активный транспорт Описывая унипорт, мы говорили об ионных каналах, но ионные каналы не единственные мембранные транспортеры в клетках. Помимо каналов существуют еще и белки-переносчики. Белки переносчики захватывают ион питательного вещества с одной стороны мембраны, а затем меняют форму и «выталкивают» ион с другой. Ну а ионные каналы это эмм… каналы. Они могут открываться и закрываться и ионы спокойно проходят через них, когда они открыты. Разница не настолько мала, как может показаться: белки-переносчики быстро достигают максимума скорости транспорта при достижении определенной концентрации – они физически не могут транспортировать больше ионов, в то время как зависимость транспорта через каналы от концентрации более ярко выражена. Белки-переносчики дают больший контроль над протеканием транспорта, а ионные каналы – большую скорость: проницаемость канала на три порядка выше, чем белка-переносчика и на 11 порядков выше, чем простая диффузия через мембрану. Наконец, мы переходим к активному транспорту. Как мы помним, активный транспорт – это перемещение ионов против электрохимического градиента, то есть туда, где их уже больше. Поверхностное представление о физике дает нам понимание, что такие вещи не могут происходить бесплатно. Так и есть – на осуществление активного транспорта требуется энергия, которую растение получает из распада молекулы АТФ – аденозин-три-фосфата. Активный транспорт делится на первично-активный и вторично-активный. Разницу обсудим позже, а пока поговорим о первично-активном транспорте В основе первично-активного транспорта лежат ионные помпы. Ионные помпы – это белки, которые связывая различные ионы, способны выталкивать их в разных направлениях независимо от градиента электрохимического потенциала. По сути это насосы для катионов. Их еще называют АТФ-азами, по запускающей их работу молекуле. Давайте подробнее рассмотрим, как работают ионные помпы на примере протонной помпы. Ион водорода H+ (гидроксоний) внутри клетки закрепляется на внутренней стороне помпы. Молекула АТФ прикрепляется к рецепторам на помпе внутри клетки и распадается. Высвобожденная энергия изменяет форму белка таким образом, что гидроксоний оказывается снаружи клетки. Как только ион водорода отсоединяется от помпы, она возвращается в изначальное состояние. Вы спросите, зачем вообще клетка выпускает ионы водорода в окружающую среду? А дело в том, что это – основной механизм получения питательных веществ растениями. Выпуская кучу ионов вплотную к клеточной стенке, растение уменьшает концентрацию других ионов возле нее, то есть как бы разбавляет их своими ионами водорода. Это создает градиент электрохимического потенциала по направлению к клетке, где потом они захватываются средствами вторичного активного транспорта, но об этом чуть ниже, а пока о помпах. Кальциевые помпы (или Ca2+-АТФазы). Клетка может выпускать ионы кальция в окружающий раствор помпами, а потом забирать их через каналы. Существует целая система активного транспорта ионов кальция, и отнюдь не только потому, что растение в нем очень нуждается как в строительном материале. Кальций – основной элемент вторичной сигнальной системы растения. Про это можно отдельную статью написать, но сейчас я расскажу вкратце. Клетки способны способны улавлить мельчайшие изменения концентрации ионов кальция, а также кальций легко взаимодействует с белками. Это связано с электрохимическими свойствами кальция. Кальциевые помпы закачивают и выкачивают кальций из окружающей среды и складов кальция внутри клетки (которые называются эндоплазматические ретикулумы, но можете развидеть это), удерживая низкий, но контролируемый уровень иона в цитоплазме, тем самым контролируя скорость протекания белковых реакций. Тяжелые металлы (такие как Fe, Cu, Mn, Zn и Mo) транспортируются в клетку большой группой транспортеров, известной как P1B. Так как все эти металлы могут быть токсичны в больших количествах, растение активно регулирует их содержание в клеточной жидкости, выталкивая излишки наружу. Все перечисленные выше АТФ-азы относятся к АТФ-азам P-типа (P здесь значит фосфор – по названию активирующего их работу элемента). Помимо них есть АТФ-азы V-типа и F-типа, первые используются для переноса веществ внутри клетки, а вторые для производства фотосинтеза. Но нам они не интересны, по крайней мере в рамках этой статьи. Отдельной группой стоят так называемые ABC-переносчики. Это огромное семейство транспортеров, которые обеспечивают мембранный транспорт пептидов, аминокислот, сахаров, липидов, ионов металлов, полисахаридов, вторичных соединений, неорганических кислот и других молекул. ABC-белки могут транспортировать фенольные соединения и алкалоиды, терпеноиды и воска, стероиды и продукты распада хлорофиллов, гербициды и тяжелые металлы. АВС-переносчики участвуют в полярном транспорте ауксина, катаболизме липидов, устойчивости к болезням, работе устьиц и детоксикации. Короче, это один из главных регуляторных механизмов растения, особенно тех, что касаются сложных органических и неорганических молекул. Тем не менее эти белки не имеют особо важной роли в питании растения, их функции другие. Отличительной особенностью их от переносчиков P-типа является наличие двух дополнительных частей (ABC-доменов) внутри клетки, что делает механизм их работы слегка другим. В открытом состоянии переносчик связывает специфичную для него молекулу, затем он закрывается, и к ABC-доменам присоединяются две молекулы АТФ, что приводит к видоизменению белка и выбросу связанной молекулы наружу. Вторично-активный транспорт Теперь, когда самое сложное уже позади и вы, я надеюсь, прочитали и даже поняли предыдущий текст, я расскажу, зачем растению нужна протонная помпа. Дело в том, что выкачать из клетки ион водорода оказывается легче и менее энергозатратно, чем транспортировать более тяжелые элементы внутрь. Поэтому клетка, выбрасывая катион водорода в околоклеточный раствор, создает электрохимический градиент этого катиона по направлению внутрь клетки. Я вкратце говорил, что такое электрохимический градиент вначале статьи, сейчас хочу рассказать подробнее. Электрохимический градиент – это по сути сумма электрического и химического градиента. Он выражается в количестве энергии, которую нужно приложить, чтобы передвинуть ион против градиента или количество энергии, которое высвобождается при движении иона по градиенту. Согласно закону энтропии, энергия стремится уравновесить себя в пространстве. Чем больше концентрация веществ с одной или другой части сообщающихся сосудов, тем больше их суммарная энергия. Поэтому ионы всегда двигаются в сторону меньшей концентрации. Это химический градиент. С другой стороны, заряженные частицы создают электрическое поле. Если в одном растворе положительно заряженных частиц больше чем в другом, то они будут перемещаться вдоль создаваемого ими магнитного поля до тех пор, пока заряды не уравновесят друг друга. Это электрический градиент. Электрохимический градиент – это сумма химического и электрического градиента. Так вот, вторично-активный транспорт для протонных помп выполняют белки-переносчики, которые являются симпортами (или котранспортерами). Помните, ранее я рассказывал, что мембранные транспортеры делятся на унипорты, симпорты и антипорты? Так вот, симпорт двигает два иона внутрь клетки, используя электрохимический градиент одного из них. Вот как это происходит: протонная помпа создает электрохимический градиент, активно выкачивая катионы водорода из клетки. Катион питательного вещества присоединяется к белку переносчику. Затем, к нему же присоединяется катион водорода. Энергия, высвобождаемая прохождением катиона водорода внутрь клетки, используется для продвижения внутрь клетки иона питательного вещества против его градиента концентрации. Как видите, энергия на активный транспорт питательного вещества затрачивается не напрямую, а опосредованно протонной помпой. Другой тип белков вторичного активного транспорты – это антипорты (или обменники). Суть та же, что и в симпортах, только в антипортах молекулы транспортируются в разных направлениях. В связке с протонной помпой и электрохимическим градиентом H+ антипорты чаще служат для выталкивания элементов наружу, а не внутрь клетки. Учитывая важность протонный помпы для жизнедеятельности растения, вы должны понимать важность pH раствора, который как я надеюсь вы знаете, является мерой концентрация катиона водорода H+. При слишком высоком pH, то есть малой концентрации катионов водорода, энергии электрохимического градиента может не хватить для переноса вещества в клетку средствами вторично-активного транспорта, а при слишком низком pH, то есть высокой концентрации Н+, катионы водорода могут забивать трансмембранны поры. Мембранный транспорт большинства катионов у растений осуществляется за счет энергии электрохимического градиента ионов H+. Таким путем в растительную клетку или из нее могут транспортироваться такие катионы, как K+, NH4+ Na+, Ca2+ и Mg2+. Белки-переносчики калия – это три группы, два симпорта и один антипорт (HKT, KT/KUP/HAK и CPA соответственно, если захотите узнать о них поболее). Антипорт калия менее специфичен, чем симпорты, через него могут выталкиваться и другие протоны, но это то – что растению и нужно – выталкивать излишки. Аммоний транспортируется переносчиками симпортами класса AMT. Их активность пропорциональна электрохимическому градиенту Н+. Помимо первично-активных кальциевых помп, растение избавляется от излишков кальция в цитоплазме с помощью Н+/Са2+ -обменников. Натрий, который не слишком нужен растению, транспортируется похожим H+/Na+ -антипортом. Транспорт магния осуществляется магниевыми помпами и обменниками, и в целом похож на транспорт кальция. Большая часть информации о типах мембранных транспортеров появилась из расшифровки генома растений и принадлежит последним двум десятилетиям. Я хочу сказать, что вероятнее всего мы еще не знаем о существовании многих видов транспортных каналов. Транспорт аниов протекает в основном симпортом с протоном водорода. Таким путем в растительную клетку поступают фосфат, нитрат и сульфат. Переносчик фосфора в плазматической мембране – это белки семейства Pht1. Pht1. Нитраты переносят NRT1 и NRT2 –симпортеры, сильно различающиеся скоростью и объемом транспорта. Сульфатные переносчики состоят из 3 типов, два из которых отвечают за переносом вещества в корневой, а третий – через лист во флоэму. Ионные каналы Вернемся к нашим каналам. Вкратце я о них уже рассказывал, теперь поподробней. Как вы уже, наверное, поняли, ионные каналы — это основной механизм пассивного транспорта в клетке – облегчённой диффузии. Избирательность канала обусловлена формой его селективного фильтра – через один вид канала может проходить один (хотя иногда и несколько) вид иона. Но каналы еще должны уметь открываться и закрываться, независимо от присутствия нужного иона, иначе их поток будет неуправляем. Положение открытия/закрытия канала (воротный механизм) регулируется набором факторов, зачастую несколькими из них. Белок может открыться из-за присоединившейся к сенсору канала молекулы (лиганда). Лиганд, присоединяясь к каналу, меняет его форму, и он открывается. Канал также может открываться из-за величины мембранного потенциала, то есть электрическому напряжению мембраны, из-за pH, растяжения мембраны и света, последнее относится к клеткам надземной части. Выход иона из канала облегчается при появлении на входе другого иона из-за их электростатического отталкивания. Однако при высоких концентрациях может происходить насыщение канала из-за заполнения ионами его входа и выхода и, как следствие, его блокировка. Через селективные каналы мембраны могут проходить в обоих направлениях ионы К+, Са2+, в меньшей степени NH4+, Ba2+, Mg2+. Но если все бы было так просто мы бы никогда не смогли бы получить в растениях токсичности по определенным элементам. Помимо селективных в клеточной мембране растений также имеются неселективные каналы. Они способны пропускать Na+, NH4+, Mg2+, Zn2+, Li+, Cs+, Ba2+ и другие катионы. Анионные каналы работают также, как и катионные, с небольшим отличием в том, что на вход им нужно преодолевать электрический потенциал, созданный протонный помпой. Поэтому анионные каналы работают только как резервные средства при сильном дефиците анионов в растении. Так их основная функция – избавление от анионов при их переизбытке в цитоплазме. Но анионные каналы существуют не только для минеральных питательных веществ. Они могут выпускать малат и цитрат, продукты метаболизма растения, которые, являясь слабыми кислотами, контролируют уровень pH вокруг корня. Еще они выпускают органические соединения, которые связывают Al3+, токсичный для многих процессов растения. Выводы На основе этой информации вы можете сделать свои выводы. Вот вам мои. Растение обладает огромными возможностями в контроле поступаемых в него питательных веществ. Это – результат эволюции в миллиарды лет. Любое вещество, попавшее в клетку растения, может быть вытолкнуто наружу. Что это значит? Что не существует никакой идеальной формулы питательного раствора, существует оптимальный диапазон для каждого питательного вещества. Идеальный раствор – это тот, в котором растение затрачивает минимум энергии на получение питательных веществ и минимум энергии на то, чтобы избавляться от питательных веществ, выбрасывая их в околоклеточный раствор. Можем ли мы высчитать объем необходимого питания используя информацию о мембранных каналах? К сожалению, нет. Во-первых, механизм работы многих транспортеров до сих пор недостаточно понятен. Во-вторых, точную информацию о количестве и типах, присутствующих в растении каналов, мы можем собрать, только лишь расшифровав геном растения. Геном каннабиса не расшифрован. Большая часть информации в этой статье построена на физиологии совсем другого растения, этакой лабораторной мыши для ботаников – резуховидки Таля, также известной как арабидопсис. Но кое-что мы все же определить можем. pH среды важен настолько, насколько он важен в прикорневой области, ризосфере. Растение может легко регулировать pH прикорневой в субстратах типа земли или кокоса. В гидропонных системах типа ДВЦ, в которых растение продолжительное время находится в контакте с раствором, регуляция pH в основном ложится в руки гровера. Общая концентрация питательных веществ важна не меньше, чем их соотношение. При высокой концентрации растению становится труднее пользоваться ионной помпой, так как оно удерживает мембранный потенциал на определенном уровне. С другой стороны, высокая концентрация благоприятно сказывается на пассивном притоке питательных веществ в растение, хотя и ухудшает работу каналов «на выход». Слишком низкая концентрация питательных веществ может сказаться еще хуже, чем их избыток – растение останавливает рост. По моему мнению легкий передоз лучше, чем легкий дефицит. Но самый лучший вариант – держать концентрацию оптимальной. Что можно сказать о соотношении элементов? Я бы разделил все питательные элементы на две группы – те, которые растение привыкло видеть в избытке и те, которые оно привыкло видеть в дефиците. К первым я бы отнес калий, кальций, магний и серу. Это те элементы, которые находятся в избытке в большинстве почв. Калий – это единственный элемент растения, который может находиться в равновесной концентрации в клетке и окружающем растворе. Растению нужно много калия, его избыток может привести к проблемам электрохимического потенциала мембраны, но сам он совершенно не токсичен для растения. Растение обладает развитой системой регулировки содержания кальция в цитоплазме. Разница концентраций кальция внутри клетки и снаружи может составлять несколько порядков. Магниевые транспортеры во многом схожи с кальциевыми, а сера – это анион, и растение может легко избавиться от нее открыв анионные каналы и выбросив в ее в слегка положительно заряженную окружающую среду. Первая группа элементов это та, на которую я бы советовал обращать меньше внимания. Растение обладает механизмами, позволяющими регулировать их абсорбацию, главное, чтобы их было достаточно. Другое дело вторая группа, к которой я отношу азот и фосфор. Азот медленно восстанавливается в почве, он базовый элемент растения в накоплении биомассы и растение привыкло забирать его полностью. Несколько видов азота в растворе позволяет задействовать больше каналов и улучшить его абсорбацию и регуляцию. С фосфором проще, так как он анион, растению легче от него избавиться. Но я считаю, что растение не могло эволюционно развить высокоэффективные механизмы регулирования его содержания в клетке. Что я хочу сказать, так это то, что соотношение питательных элементов в растворе можно свести в большей степени к количеству азота, в меньшей – фосфора и в совсем небольшой к количеству других макроэлементов. Что касается микроэлементов, в большой концентрации они токсичны для растения, поэтому оно обладает механизмами удержания их в клетке в необходимой концентрации, даже когда разница концентрации с околоклеточным раствором составляет несколько порядков. Я бы советовал обращать внимание только на бор, который может поступать через аквапорины, хлор и натрий, присутствие которых в субстратах может быть чрезмерным для компенсаторных механизмов растения. Лучшей инструкцией как правильно кормить является ваш опыт и интуиция, которые вы должны «прокачать», прочитав эту статью. Следите за азотом, pH, и общей концентрацией. Спасибо всем, кто дочитал эту простыню до конца. Удачных гровов и больших шишек! Аминь! Статья победитель апрельского конкурса "Автор, жги" Читай также: Гроупедия - огромная энциклопедия по выращиванию. Постоянно пополняется интересными материалами. Гид по поливу растений Анионы и катионы: сбалансирована ли ваша система?
Хочу сразу предупредить особо рьяных критиков: в данном теме я не рассматриваю эндо- и экзоцитоз, то есть регуляцию растениями цельных незаряженных молекул питательных веществ. В какой-то мере по этой причине информация здесь может считаться неполной, но тем не менее, я считаю, что данный текст будет полезен тем, кто хочет прикоснуться к настоящей научной агрономии. Мембранный транспорт Мембранный транспорт – это общее название всех процессов транспортировки веществ в клетку и из клетки через клеточную мембрану. Мембраны обладают избирательной проницаемостью, что означает, что только определенные молекулы и частицы способны проникать через нее. Клеточная мембрана устроена намного сложнее, чем любая искусственная мембрана, в первую очередь потому, что мембранный транспорт клетки обеспечивают белки переносчики, которые изменяют проницаемость мембраны в сторону большей или меньшей проницаемости для определенных растворов. К сожалению, это не означает, что растение может самостоятельно регулировать проницаемость клеточной мембраны – только то, что проницаемость мембраны может быть разная даже в похожих клетках. Мембранный транспорт может осуществляться по или против градиента электрохимического потенциала. Электрохимический потенциал можно представить, как давление на клеточную стенку либо изнутри, либо снаружи, но не физическое давление, как давление воды на плотину, а как притяжение заряженных частиц к одной из сторон. Движение к более низкому электрохимическому потенциалу не требует затрат энергии, и называется пассивным транспортом. Движение против градиента требует затрат энергии и называется активным транспортом. Здесь очень важный момент, следует отличать градиент электрохимического потенциала от градиента концентрации (водного потенциала). В сообщающихся растворах все элементы стремятся к равной концентрации независимо от других. Градиент концентрации указывает на стремлении к равной концентрации растворителя, то есть воды. Когда мы измеряем электропроводимость раствора, мы измеряем концентрацию ионов в нем, но это не говорит о том, больше ли, например, ионов калия в одном растворе, чем в другом. Но это говорит о том где больше концентрация воды, и в какую сторону направлен ее потенциал. То есть, пользуясь кондуктометром мы на самом деле измеряем концентрацию воды. Но если в первом сосуде концентрация ионов, например, кальция будет выше, чем во втором, то ионы будут перемещаться из первого в сторону второго, независимо от градиента концентрации между растворами Пассивный транспорт Прежде чем мы рассмотрим пассивный транспорт, поговорим о том, как вообще устроена клеточная мембрана или, как она еще называется, плазмалитическая мембрана. Она представляет собой двойной слой молекул фосфолипидов, состоящих из гидрофильной (водопроницаемой) головки и гидрофобного (водонепроницаемого) хвоста. Молекулы расположены головками наружу – в клетку и окружающую ее среду, а хвосты образуют гидрофобный слой, ограждающий клетку от диффузии между клеточной жидкостью и раствором снаружи. Плотный билипидный слой прерывается белками – каналами и переносчиками, которые и обеспечивают основный обмен элементами между клеткой и окружающей средой. Осмос Для начала следует понять, как именно вода поступает в клетку растения. Вода всегда перемещается по градиенту концентрации – в тот раствор, где ее концентрация меньше. В нормальных условиях концентрация воды выше внутри клетки, чем снаружи, но как вода преодолевает гидрофобный слой плазматической мембраны? Помогают ей в этом специальные белки – аквапорины. Мономеры аквапоринов образуют водяную пору, сужающуюся структуру, через которую вода проходит как песок через песочные часы. Аквапорины не пропускают заряженные частицы – ионы питательных веществ, однако помимо воды через них могут также проходить небольшие незаряженные частицы - мочевина, глицерол, борная и кремниевая кислоты, перекись водорода, а также газы (CO2 , NH3 , NO). Читайте также: Детка, ты просто осмос! Диффузия Через мембрану клетки могут беспрепятственно проходить только очень маленькие неполярные молекулы, такие как газы O2 и CO2. Билипидный барьер надежно защищает клетку от неконтролируемого проникновения любой жидкости или питательных веществ. Тем не менее, диффузия может произойти вследствие повреждения клеточной мембраны. Но это скорее аномалия, поэтому мы ее рассматривать не будем, так как ее влияние на питание растения ничтожно. Облегчённая диффузия Облегчённая диффузия (название то какое) – это диффузия через специальные каналы, которые клетка заботливо и заблаговременно оставила в клеточной мембране. Это не просто дырки в мембране, это совершенно другой процесс. В его основе лежит унипорт. Небольшое отступление – унипорт, симпорт и антипорт – так можно разделить мембранные транспортеры по количеству проходящих через них ионов и их направлению. Унипорт – это канал, в который проходит один ион в одном направлении. Симпорт – два иона в одном и том же направлении. Антипорт – два иона одновременно проходят через канал в противоположных направлениях. Так вот, белки, через которые проходит облегченная диффузия – это унипорты. В отличии от обычной диффузии транспорт через каналы отличается избирательностью – через один вид канала может проходить только лишь определенный вид ионов. Также отличия есть и в зависимости скорости транспорта от разницы концентраций и возможности подавления работы каналов специальными ингибиторами. Активный транспорт Описывая унипорт, мы говорили об ионных каналах, но ионные каналы не единственные мембранные транспортеры в клетках. Помимо каналов существуют еще и белки-переносчики. Белки переносчики захватывают ион питательного вещества с одной стороны мембраны, а затем меняют форму и «выталкивают» ион с другой. Ну а ионные каналы это эмм… каналы. Они могут открываться и закрываться и ионы спокойно проходят через них, когда они открыты. Разница не настолько мала, как может показаться: белки-переносчики быстро достигают максимума скорости транспорта при достижении определенной концентрации – они физически не могут транспортировать больше ионов, в то время как зависимость транспорта через каналы от концентрации более ярко выражена. Белки-переносчики дают больший контроль над протеканием транспорта, а ионные каналы – большую скорость: проницаемость канала на три порядка выше, чем белка-переносчика и на 11 порядков выше, чем простая диффузия через мембрану. Наконец, мы переходим к активному транспорту. Как мы помним, активный транспорт – это перемещение ионов против электрохимического градиента, то есть туда, где их уже больше. Поверхностное представление о физике дает нам понимание, что такие вещи не могут происходить бесплатно. Так и есть – на осуществление активного транспорта требуется энергия, которую растение получает из распада молекулы АТФ – аденозин-три-фосфата. Активный транспорт делится на первично-активный и вторично-активный. Разницу обсудим позже, а пока поговорим о первично-активном транспорте В основе первично-активного транспорта лежат ионные помпы. Ионные помпы – это белки, которые связывая различные ионы, способны выталкивать их в разных направлениях независимо от градиента электрохимического потенциала. По сути это насосы для катионов. Их еще называют АТФ-азами, по запускающей их работу молекуле. Давайте подробнее рассмотрим, как работают ионные помпы на примере протонной помпы. Ион водорода H+ (гидроксоний) внутри клетки закрепляется на внутренней стороне помпы. Молекула АТФ прикрепляется к рецепторам на помпе внутри клетки и распадается. Высвобожденная энергия изменяет форму белка таким образом, что гидроксоний оказывается снаружи клетки. Как только ион водорода отсоединяется от помпы, она возвращается в изначальное состояние. Вы спросите, зачем вообще клетка выпускает ионы водорода в окружающую среду? А дело в том, что это – основной механизм получения питательных веществ растениями. Выпуская кучу ионов вплотную к клеточной стенке, растение уменьшает концентрацию других ионов возле нее, то есть как бы разбавляет их своими ионами водорода. Это создает градиент электрохимического потенциала по направлению к клетке, где потом они захватываются средствами вторичного активного транспорта, но об этом чуть ниже, а пока о помпах. Кальциевые помпы (или Ca2+-АТФазы). Клетка может выпускать ионы кальция в окружающий раствор помпами, а потом забирать их через каналы. Существует целая система активного транспорта ионов кальция, и отнюдь не только потому, что растение в нем очень нуждается как в строительном материале. Кальций – основной элемент вторичной сигнальной системы растения. Про это можно отдельную статью написать, но сейчас я расскажу вкратце. Клетки способны способны улавлить мельчайшие изменения концентрации ионов кальция, а также кальций легко взаимодействует с белками. Это связано с электрохимическими свойствами кальция. Кальциевые помпы закачивают и выкачивают кальций из окружающей среды и складов кальция внутри клетки (которые называются эндоплазматические ретикулумы, но можете развидеть это), удерживая низкий, но контролируемый уровень иона в цитоплазме, тем самым контролируя скорость протекания белковых реакций. Тяжелые металлы (такие как Fe, Cu, Mn, Zn и Mo) транспортируются в клетку большой группой транспортеров, известной как P1B. Так как все эти металлы могут быть токсичны в больших количествах, растение активно регулирует их содержание в клеточной жидкости, выталкивая излишки наружу. Все перечисленные выше АТФ-азы относятся к АТФ-азам P-типа (P здесь значит фосфор – по названию активирующего их работу элемента). Помимо них есть АТФ-азы V-типа и F-типа, первые используются для переноса веществ внутри клетки, а вторые для производства фотосинтеза. Но нам они не интересны, по крайней мере в рамках этой статьи. Отдельной группой стоят так называемые ABC-переносчики. Это огромное семейство транспортеров, которые обеспечивают мембранный транспорт пептидов, аминокислот, сахаров, липидов, ионов металлов, полисахаридов, вторичных соединений, неорганических кислот и других молекул. ABC-белки могут транспортировать фенольные соединения и алкалоиды, терпеноиды и воска, стероиды и продукты распада хлорофиллов, гербициды и тяжелые металлы. АВС-переносчики участвуют в полярном транспорте ауксина, катаболизме липидов, устойчивости к болезням, работе устьиц и детоксикации. Короче, это один из главных регуляторных механизмов растения, особенно тех, что касаются сложных органических и неорганических молекул. Тем не менее эти белки не имеют особо важной роли в питании растения, их функции другие. Отличительной особенностью их от переносчиков P-типа является наличие двух дополнительных частей (ABC-доменов) внутри клетки, что делает механизм их работы слегка другим. В открытом состоянии переносчик связывает специфичную для него молекулу, затем он закрывается, и к ABC-доменам присоединяются две молекулы АТФ, что приводит к видоизменению белка и выбросу связанной молекулы наружу. Вторично-активный транспорт Теперь, когда самое сложное уже позади и вы, я надеюсь, прочитали и даже поняли предыдущий текст, я расскажу, зачем растению нужна протонная помпа. Дело в том, что выкачать из клетки ион водорода оказывается легче и менее энергозатратно, чем транспортировать более тяжелые элементы внутрь. Поэтому клетка, выбрасывая катион водорода в околоклеточный раствор, создает электрохимический градиент этого катиона по направлению внутрь клетки. Я вкратце говорил, что такое электрохимический градиент вначале статьи, сейчас хочу рассказать подробнее. Электрохимический градиент – это по сути сумма электрического и химического градиента. Он выражается в количестве энергии, которую нужно приложить, чтобы передвинуть ион против градиента или количество энергии, которое высвобождается при движении иона по градиенту. Согласно закону энтропии, энергия стремится уравновесить себя в пространстве. Чем больше концентрация веществ с одной или другой части сообщающихся сосудов, тем больше их суммарная энергия. Поэтому ионы всегда двигаются в сторону меньшей концентрации. Это химический градиент. С другой стороны, заряженные частицы создают электрическое поле. Если в одном растворе положительно заряженных частиц больше чем в другом, то они будут перемещаться вдоль создаваемого ими магнитного поля до тех пор, пока заряды не уравновесят друг друга. Это электрический градиент. Электрохимический градиент – это сумма химического и электрического градиента. Так вот, вторично-активный транспорт для протонных помп выполняют белки-переносчики, которые являются симпортами (или котранспортерами). Помните, ранее я рассказывал, что мембранные транспортеры делятся на унипорты, симпорты и антипорты? Так вот, симпорт двигает два иона внутрь клетки, используя электрохимический градиент одного из них. Вот как это происходит: протонная помпа создает электрохимический градиент, активно выкачивая катионы водорода из клетки. Катион питательного вещества присоединяется к белку переносчику. Затем, к нему же присоединяется катион водорода. Энергия, высвобождаемая прохождением катиона водорода внутрь клетки, используется для продвижения внутрь клетки иона питательного вещества против его градиента концентрации. Как видите, энергия на активный транспорт питательного вещества затрачивается не напрямую, а опосредованно протонной помпой. Другой тип белков вторичного активного транспорты – это антипорты (или обменники). Суть та же, что и в симпортах, только в антипортах молекулы транспортируются в разных направлениях. В связке с протонной помпой и электрохимическим градиентом H+ антипорты чаще служат для выталкивания элементов наружу, а не внутрь клетки. Учитывая важность протонный помпы для жизнедеятельности растения, вы должны понимать важность pH раствора, который как я надеюсь вы знаете, является мерой концентрация катиона водорода H+. При слишком высоком pH, то есть малой концентрации катионов водорода, энергии электрохимического градиента может не хватить для переноса вещества в клетку средствами вторично-активного транспорта, а при слишком низком pH, то есть высокой концентрации Н+, катионы водорода могут забивать трансмембранны поры. Мембранный транспорт большинства катионов у растений осуществляется за счет энергии электрохимического градиента ионов H+. Таким путем в растительную клетку или из нее могут транспортироваться такие катионы, как K+, NH4+ Na+, Ca2+ и Mg2+. Белки-переносчики калия – это три группы, два симпорта и один антипорт (HKT, KT/KUP/HAK и CPA соответственно, если захотите узнать о них поболее). Антипорт калия менее специфичен, чем симпорты, через него могут выталкиваться и другие протоны, но это то – что растению и нужно – выталкивать излишки. Аммоний транспортируется переносчиками симпортами класса AMT. Их активность пропорциональна электрохимическому градиенту Н+. Помимо первично-активных кальциевых помп, растение избавляется от излишков кальция в цитоплазме с помощью Н+/Са2+ -обменников. Натрий, который не слишком нужен растению, транспортируется похожим H+/Na+ -антипортом. Транспорт магния осуществляется магниевыми помпами и обменниками, и в целом похож на транспорт кальция. Большая часть информации о типах мембранных транспортеров появилась из расшифровки генома растений и принадлежит последним двум десятилетиям. Я хочу сказать, что вероятнее всего мы еще не знаем о существовании многих видов транспортных каналов. Транспорт аниов протекает в основном симпортом с протоном водорода. Таким путем в растительную клетку поступают фосфат, нитрат и сульфат. Переносчик фосфора в плазматической мембране – это белки семейства Pht1. Pht1. Нитраты переносят NRT1 и NRT2 –симпортеры, сильно различающиеся скоростью и объемом транспорта. Сульфатные переносчики состоят из 3 типов, два из которых отвечают за переносом вещества в корневой, а третий – через лист во флоэму. Ионные каналы Вернемся к нашим каналам. Вкратце я о них уже рассказывал, теперь поподробней. Как вы уже, наверное, поняли, ионные каналы — это основной механизм пассивного транспорта в клетке – облегчённой диффузии. Избирательность канала обусловлена формой его селективного фильтра – через один вид канала может проходить один (хотя иногда и несколько) вид иона. Но каналы еще должны уметь открываться и закрываться, независимо от присутствия нужного иона, иначе их поток будет неуправляем. Положение открытия/закрытия канала (воротный механизм) регулируется набором факторов, зачастую несколькими из них. Белок может открыться из-за присоединившейся к сенсору канала молекулы (лиганда). Лиганд, присоединяясь к каналу, меняет его форму, и он открывается. Канал также может открываться из-за величины мембранного потенциала, то есть электрическому напряжению мембраны, из-за pH, растяжения мембраны и света, последнее относится к клеткам надземной части. Выход иона из канала облегчается при появлении на входе другого иона из-за их электростатического отталкивания. Однако при высоких концентрациях может происходить насыщение канала из-за заполнения ионами его входа и выхода и, как следствие, его блокировка. Через селективные каналы мембраны могут проходить в обоих направлениях ионы К+, Са2+, в меньшей степени NH4+, Ba2+, Mg2+. Но если все бы было так просто мы бы никогда не смогли бы получить в растениях токсичности по определенным элементам. Помимо селективных в клеточной мембране растений также имеются неселективные каналы. Они способны пропускать Na+, NH4+, Mg2+, Zn2+, Li+, Cs+, Ba2+ и другие катионы. Анионные каналы работают также, как и катионные, с небольшим отличием в том, что на вход им нужно преодолевать электрический потенциал, созданный протонный помпой. Поэтому анионные каналы работают только как резервные средства при сильном дефиците анионов в растении. Так их основная функция – избавление от анионов при их переизбытке в цитоплазме. Но анионные каналы существуют не только для минеральных питательных веществ. Они могут выпускать малат и цитрат, продукты метаболизма растения, которые, являясь слабыми кислотами, контролируют уровень pH вокруг корня. Еще они выпускают органические соединения, которые связывают Al3+, токсичный для многих процессов растения. Выводы На основе этой информации вы можете сделать свои выводы. Вот вам мои. Растение обладает огромными возможностями в контроле поступаемых в него питательных веществ. Это – результат эволюции в миллиарды лет. Любое вещество, попавшее в клетку растения, может быть вытолкнуто наружу. Что это значит? Что не существует никакой идеальной формулы питательного раствора, существует оптимальный диапазон для каждого питательного вещества. Идеальный раствор – это тот, в котором растение затрачивает минимум энергии на получение питательных веществ и минимум энергии на то, чтобы избавляться от питательных веществ, выбрасывая их в околоклеточный раствор. Можем ли мы высчитать объем необходимого питания используя информацию о мембранных каналах? К сожалению, нет. Во-первых, механизм работы многих транспортеров до сих пор недостаточно понятен. Во-вторых, точную информацию о количестве и типах, присутствующих в растении каналов, мы можем собрать, только лишь расшифровав геном растения. Геном каннабиса не расшифрован. Большая часть информации в этой статье построена на физиологии совсем другого растения, этакой лабораторной мыши для ботаников – резуховидки Таля, также известной как арабидопсис. Но кое-что мы все же определить можем. pH среды важен настолько, насколько он важен в прикорневой области, ризосфере. Растение может легко регулировать pH прикорневой в субстратах типа земли или кокоса. В гидропонных системах типа ДВЦ, в которых растение продолжительное время находится в контакте с раствором, регуляция pH в основном ложится в руки гровера. Общая концентрация питательных веществ важна не меньше, чем их соотношение. При высокой концентрации растению становится труднее пользоваться ионной помпой, так как оно удерживает мембранный потенциал на определенном уровне. С другой стороны, высокая концентрация благоприятно сказывается на пассивном притоке питательных веществ в растение, хотя и ухудшает работу каналов «на выход». Слишком низкая концентрация питательных веществ может сказаться еще хуже, чем их избыток – растение останавливает рост. По моему мнению легкий передоз лучше, чем легкий дефицит. Но самый лучший вариант – держать концентрацию оптимальной. Что можно сказать о соотношении элементов? Я бы разделил все питательные элементы на две группы – те, которые растение привыкло видеть в избытке и те, которые оно привыкло видеть в дефиците. К первым я бы отнес калий, кальций, магний и серу. Это те элементы, которые находятся в избытке в большинстве почв. Калий – это единственный элемент растения, который может находиться в равновесной концентрации в клетке и окружающем растворе. Растению нужно много калия, его избыток может привести к проблемам электрохимического потенциала мембраны, но сам он совершенно не токсичен для растения. Растение обладает развитой системой регулировки содержания кальция в цитоплазме. Разница концентраций кальция внутри клетки и снаружи может составлять несколько порядков. Магниевые транспортеры во многом схожи с кальциевыми, а сера – это анион, и растение может легко избавиться от нее открыв анионные каналы и выбросив в ее в слегка положительно заряженную окружающую среду. Первая группа элементов это та, на которую я бы советовал обращать меньше внимания. Растение обладает механизмами, позволяющими регулировать их абсорбацию, главное, чтобы их было достаточно. Другое дело вторая группа, к которой я отношу азот и фосфор. Азот медленно восстанавливается в почве, он базовый элемент растения в накоплении биомассы и растение привыкло забирать его полностью. Несколько видов азота в растворе позволяет задействовать больше каналов и улучшить его абсорбацию и регуляцию. С фосфором проще, так как он анион, растению легче от него избавиться. Но я считаю, что растение не могло эволюционно развить высокоэффективные механизмы регулирования его содержания в клетке. Что я хочу сказать, так это то, что соотношение питательных элементов в растворе можно свести в большей степени к количеству азота, в меньшей – фосфора и в совсем небольшой к количеству других макроэлементов. Что касается микроэлементов, в большой концентрации они токсичны для растения, поэтому оно обладает механизмами удержания их в клетке в необходимой концентрации, даже когда разница концентрации с околоклеточным раствором составляет несколько порядков. Я бы советовал обращать внимание только на бор, который может поступать через аквапорины, хлор и натрий, присутствие которых в субстратах может быть чрезмерным для компенсаторных механизмов растения. Лучшей инструкцией как правильно кормить является ваш опыт и интуиция, которые вы должны «прокачать», прочитав эту статью. Следите за азотом, pH, и общей концентрацией. Спасибо всем, кто дочитал эту простыню до конца. Удачных гровов и больших шишек! Аминь! Статья победитель апрельского конкурса "Автор, жги" Читай также: Гроупедия - огромная энциклопедия по выращиванию. Постоянно пополняется интересными материалами. Гид по поливу растений Анионы и катионы: сбалансирована ли ваша система?- 12 комментариев
-
- 1
-
-
- физиология растений
- мембрана
- (и ещё 3 )
-
Эта книга - целый курс анатомии растений. Подойдет для тех, кто мечтал стать профессором ботаники, но что то пошло не так, ну и для гуру грова. В учебнике отражены современные представления по основным направлениям физиологии растений — фотосинтезу, дыханию, водному обмену, минеральному питанию, мембранному и дальнему транспорту веществ, фитогормонам, росту и развитию, размножению растений, устойчивости и адаптации к неблагоприятным факторам среды и патогенам, вторичному метаболизму растений, системам регуляции физиологических процессов. В основу учебника положен общий курс лекций "Физиология и биохимия растений", читаемый автором для студентов биологопочвенного факультета Санкт-Петербургского государственного университета. Скачать
- 12 ответов
-
- 9
-
-
- физиология растений
- ботаника
- (и ещё 1 )
-
 В учебнике отражены современные представления по основным направлениям физиологии растений — фотосинтезу, дыханию, водному обмену, минеральному питанию, мембранному и дальнему транспорту веществ, фитогормонам, росту и развитию, размножению растений, устойчивости и адаптации к неблагоприятным факторам среды и патогенам, вторичному метаболизму растений, системам регуляции физиологических процессов. В основу учебника положен общий курс лекций "Физиология и биохимия растений", читаемый автором для студентов биологопочвенного факультета Санкт-Петербургского государственного университета. Скачать
В учебнике отражены современные представления по основным направлениям физиологии растений — фотосинтезу, дыханию, водному обмену, минеральному питанию, мембранному и дальнему транспорту веществ, фитогормонам, росту и развитию, размножению растений, устойчивости и адаптации к неблагоприятным факторам среды и патогенам, вторичному метаболизму растений, системам регуляции физиологических процессов. В основу учебника положен общий курс лекций "Физиология и биохимия растений", читаемый автором для студентов биологопочвенного факультета Санкт-Петербургского государственного университета. Скачать -
 Агротехника-система приемов возделывания культурных растений Например на кусте обычного синего баклажана оставляют четыре баклажана. Остальные завязи удаляют. Тогда баклажаны получаются не слишком большие и не слишком мелкие, как если бы оставили три или пять. Дыню в холодном климате оставляют одну и подвешивают в мешочке над землей. Она не должна касаться земли, чтобы в холоде не загнить и не должна оторваться от плети, когда вырастет. А если пожадничать, и оставить две завязи - вы не получите ни одной дыни, так как то, что вырастет, вы не будете есть, один корень две дыни не вывезет. Наше растение Оно однолетнее. Его можно долго держать на веге, но выращивают его как однолетнее. Оно не делает запасов на следующий год в виде кочерыжки или луковицы, т.е. переводя световой режим на цветение, вы включаете ему обратный отсчет. Оно умрёт, сенсимилья (в природе семена) его высосет и убьет в итоге. Самое слабое звено у нашего растения - это корни. Они всегда снабжаются по остаточному принципу. Им нужно расти весь цикл: сначала на корне появляются корневые волоски (только они могут всасывать ионы вместе с водой). Средний срок их жизни - три дня, потом большинство отмирает, и только очень некоторые покрываются лубом и становятся настоящими корнями (волоски должны продолжать расти всё время). Ионы, попавшие в корень, дальше двигаются по транспортной системе вверх. У растения транспортных систем две: одна сеть сосудов расположена под корой, это лыко или волокно конопли называется ксилема. Она состоит из трубочек, эти трубочки состоят из одеревенелых клеток, по ним ионы двигаются вверх вместе с восходящим потоком сока. Сок этот двигают, так называемые, концевые двигатели, верхний и нижний. Подробно про работу нижнего не буду, если растение срубить - выделится на срубе жидкость – пасока. Это работа нижнего концевого двигателя, он слабее верхнего. Верхний концевой двигатель работает за счёт испарения воды через устьица - транспирации и выделения воды из желёзок по краю листа-гуттации. Второй пучок волокон расположен внутри стебля, называется флоэма. По ней вещества двигаются сверху к корням. Она состоит из живых клеток, по ней сок не двигается, по ней перемещаются вещества с затратой энергии. Вообще ксилема считается проводником ионов, а флоэма органических веществ, но по ксилеме могут двигаться аминокислоты и по флоэме калий, фосфор, а так же между ними есть сообщение - некоторые вещества могут перемещаться радиально, из флоэмы в ксилему и наоборот. Это явление называется транслокацией (это выгодно некоторым веществам - не надо делать круг по растению). Ионы из корня попадают по ксилеме в листья и меристемы надземной части. Меристемы - это область усиленного деления клеток, где происходит основной рост массы. На веге - это точки роста и молодые листья, на цветении - это шишки (там больше всего гормонов, клетки быстрее всего делятся, там больше всего расходуется продукция фотосинтеза). И только во вторую очередь снабжаются по флоэме питанием меристемы корня, при этом еще энергию надо тратить, то есть, опять же продукцию фотосинтеза. Продуктивность фотосинтеза Это самое главное. Здоровая корневая система + правильное питание удобрениями, толстая у основания кочерыжка (хорошо развитая транспортная система) , правильный климат + хорошее освещение + дополнительная подача СО2 – всё это делается, чтобы превратить углекислый газ и воду с помощью света в одно вкусное, но не полезное для человека в чистом виде вещество. Чем больше в граммах сахарозы вы получите за цикл, тем на большее количество шишек вправе рассчитывать. Из сахарозы в растении делаются все остальные органические и пластические вещества, из которых лепятся новые ткани растения и берётся энергия для энергозатратных процессов. Зачем можно отрывать листья? 1. Сформировать толстый ствол, остановить рост вверх. Но есть несколько менее варварских способов это сделать, можно прочитать например здесь (знание английского обязательно) 2. Смоделировать нападение насекомых или животного, заставить растение выработать иммунитет и выделить еще смолы. Но есть препараты с действием более сильным, безвредные для человека и оставляющие листья в покое, на основе хитозана. Bud Factor X, например. Почему нельзя отрывать листья? Что происходит, когда вы обрываете листья? Растение выделяет гормоны старения и стресса - этилен и абсцизовую кислоту. Растение под действием абсцизовой кислоты закрывает устьица и прекращает транспирацию и фотосинтез. Если сильно оборвали – на несколько дней свет будет пропадать зря. Растение под действием этилена будет наращивать ствол и перераспределять токи в изменившемся объеме. Если оборвёте очень много - этилен состарит нижние листья, а абсцизовая кислота заставит их отвалиться. Растение ре-утилизирует органические вещества из этих листьев, фотосинтеза-то нет на попытку отрастить новые листья. А так как основную массу органических веществ отнимает меристема, в условиях дефицита оных у вас рвётся самое слабое звено – корни (им ничего не достается). Корневые волоски надолго перестают расти. Дальше вы ломаете верхний концевой двигатель, при том, что нижний двигатель вы тоже уже сломали, гуттация и транспирация в прежнем объеме восстановится, когда новые листья отрастут (если отрастут). Дальше. Все листья на растении разные: верхние с толстым защитным слоем из воска-кутикулы настроены на сильный свет и предохраняют другие, те, что в глубине куста от ожогов, их хлоропласты настроены собирать слабый рассеянный свет (прямой их обжигает). Дальше . Фоточувствительность. Растение эволюционировало на одном месте, куда бы семечко не попало - в кусты или другую тень, у него есть механизм, как расположить листовую мозаику так, чтобы из любой ситуации выжать максимум света, поворотный механизм, фототропизм. Самые фоточувствительные лопухи у ствола. Очень чувствительный механизм – наш глаз, еле различает день-ночь по сравнению с ним. Если вы считаете, что куст что-то неправильно освещает, то нет. Дальше. Вы лишаетесь сахара. Не только того, который лист производит, но и того, который растение могло бы взять на ре-утилизацию. Для повторного использования могут использоваться и некоторые ионы, т.н. подвижные, чей недостаток проявляется на старых листьях. Ими растение жертвует, чтобы восполнить недостаток - молибден, магний, азот и т.д. Их запас вы тоже теряете. Дальше. Затрудняется транслокация. Она происходит в листьях, где флоэма и ксилема расположены ближе всего - в жилке листа. Дальше вы просто можете заразить растение через множество открытых ран. Заключение Наше растение по агротехнике относится к растениям, у которых удаляются лишние точки цветения. Если на предцвете в низу или внутри куста из пазухи листа лезет зародыш шишки на длинной палочке - их стоит удалить, из них путного ничего не будет, даже если дать близлежащим листьям свет (цветам свет вообще не нужен). Формировать куст с как можно более короткими расстояниями между нодами (узлами), по удочкам тяжело вещества гонять, отсекать затененные ветви, они потом начнут атрофироваться, если совсем в тень попадут. Сразу замазывать раны антисептиком для ран на растениях - Садовым Варом или РанНет. Обсудить на форуме
Агротехника-система приемов возделывания культурных растений Например на кусте обычного синего баклажана оставляют четыре баклажана. Остальные завязи удаляют. Тогда баклажаны получаются не слишком большие и не слишком мелкие, как если бы оставили три или пять. Дыню в холодном климате оставляют одну и подвешивают в мешочке над землей. Она не должна касаться земли, чтобы в холоде не загнить и не должна оторваться от плети, когда вырастет. А если пожадничать, и оставить две завязи - вы не получите ни одной дыни, так как то, что вырастет, вы не будете есть, один корень две дыни не вывезет. Наше растение Оно однолетнее. Его можно долго держать на веге, но выращивают его как однолетнее. Оно не делает запасов на следующий год в виде кочерыжки или луковицы, т.е. переводя световой режим на цветение, вы включаете ему обратный отсчет. Оно умрёт, сенсимилья (в природе семена) его высосет и убьет в итоге. Самое слабое звено у нашего растения - это корни. Они всегда снабжаются по остаточному принципу. Им нужно расти весь цикл: сначала на корне появляются корневые волоски (только они могут всасывать ионы вместе с водой). Средний срок их жизни - три дня, потом большинство отмирает, и только очень некоторые покрываются лубом и становятся настоящими корнями (волоски должны продолжать расти всё время). Ионы, попавшие в корень, дальше двигаются по транспортной системе вверх. У растения транспортных систем две: одна сеть сосудов расположена под корой, это лыко или волокно конопли называется ксилема. Она состоит из трубочек, эти трубочки состоят из одеревенелых клеток, по ним ионы двигаются вверх вместе с восходящим потоком сока. Сок этот двигают, так называемые, концевые двигатели, верхний и нижний. Подробно про работу нижнего не буду, если растение срубить - выделится на срубе жидкость – пасока. Это работа нижнего концевого двигателя, он слабее верхнего. Верхний концевой двигатель работает за счёт испарения воды через устьица - транспирации и выделения воды из желёзок по краю листа-гуттации. Второй пучок волокон расположен внутри стебля, называется флоэма. По ней вещества двигаются сверху к корням. Она состоит из живых клеток, по ней сок не двигается, по ней перемещаются вещества с затратой энергии. Вообще ксилема считается проводником ионов, а флоэма органических веществ, но по ксилеме могут двигаться аминокислоты и по флоэме калий, фосфор, а так же между ними есть сообщение - некоторые вещества могут перемещаться радиально, из флоэмы в ксилему и наоборот. Это явление называется транслокацией (это выгодно некоторым веществам - не надо делать круг по растению). Ионы из корня попадают по ксилеме в листья и меристемы надземной части. Меристемы - это область усиленного деления клеток, где происходит основной рост массы. На веге - это точки роста и молодые листья, на цветении - это шишки (там больше всего гормонов, клетки быстрее всего делятся, там больше всего расходуется продукция фотосинтеза). И только во вторую очередь снабжаются по флоэме питанием меристемы корня, при этом еще энергию надо тратить, то есть, опять же продукцию фотосинтеза. Продуктивность фотосинтеза Это самое главное. Здоровая корневая система + правильное питание удобрениями, толстая у основания кочерыжка (хорошо развитая транспортная система) , правильный климат + хорошее освещение + дополнительная подача СО2 – всё это делается, чтобы превратить углекислый газ и воду с помощью света в одно вкусное, но не полезное для человека в чистом виде вещество. Чем больше в граммах сахарозы вы получите за цикл, тем на большее количество шишек вправе рассчитывать. Из сахарозы в растении делаются все остальные органические и пластические вещества, из которых лепятся новые ткани растения и берётся энергия для энергозатратных процессов. Зачем можно отрывать листья? 1. Сформировать толстый ствол, остановить рост вверх. Но есть несколько менее варварских способов это сделать, можно прочитать например здесь (знание английского обязательно) 2. Смоделировать нападение насекомых или животного, заставить растение выработать иммунитет и выделить еще смолы. Но есть препараты с действием более сильным, безвредные для человека и оставляющие листья в покое, на основе хитозана. Bud Factor X, например. Почему нельзя отрывать листья? Что происходит, когда вы обрываете листья? Растение выделяет гормоны старения и стресса - этилен и абсцизовую кислоту. Растение под действием абсцизовой кислоты закрывает устьица и прекращает транспирацию и фотосинтез. Если сильно оборвали – на несколько дней свет будет пропадать зря. Растение под действием этилена будет наращивать ствол и перераспределять токи в изменившемся объеме. Если оборвёте очень много - этилен состарит нижние листья, а абсцизовая кислота заставит их отвалиться. Растение ре-утилизирует органические вещества из этих листьев, фотосинтеза-то нет на попытку отрастить новые листья. А так как основную массу органических веществ отнимает меристема, в условиях дефицита оных у вас рвётся самое слабое звено – корни (им ничего не достается). Корневые волоски надолго перестают расти. Дальше вы ломаете верхний концевой двигатель, при том, что нижний двигатель вы тоже уже сломали, гуттация и транспирация в прежнем объеме восстановится, когда новые листья отрастут (если отрастут). Дальше. Все листья на растении разные: верхние с толстым защитным слоем из воска-кутикулы настроены на сильный свет и предохраняют другие, те, что в глубине куста от ожогов, их хлоропласты настроены собирать слабый рассеянный свет (прямой их обжигает). Дальше . Фоточувствительность. Растение эволюционировало на одном месте, куда бы семечко не попало - в кусты или другую тень, у него есть механизм, как расположить листовую мозаику так, чтобы из любой ситуации выжать максимум света, поворотный механизм, фототропизм. Самые фоточувствительные лопухи у ствола. Очень чувствительный механизм – наш глаз, еле различает день-ночь по сравнению с ним. Если вы считаете, что куст что-то неправильно освещает, то нет. Дальше. Вы лишаетесь сахара. Не только того, который лист производит, но и того, который растение могло бы взять на ре-утилизацию. Для повторного использования могут использоваться и некоторые ионы, т.н. подвижные, чей недостаток проявляется на старых листьях. Ими растение жертвует, чтобы восполнить недостаток - молибден, магний, азот и т.д. Их запас вы тоже теряете. Дальше. Затрудняется транслокация. Она происходит в листьях, где флоэма и ксилема расположены ближе всего - в жилке листа. Дальше вы просто можете заразить растение через множество открытых ран. Заключение Наше растение по агротехнике относится к растениям, у которых удаляются лишние точки цветения. Если на предцвете в низу или внутри куста из пазухи листа лезет зародыш шишки на длинной палочке - их стоит удалить, из них путного ничего не будет, даже если дать близлежащим листьям свет (цветам свет вообще не нужен). Формировать куст с как можно более короткими расстояниями между нодами (узлами), по удочкам тяжело вещества гонять, отсекать затененные ветви, они потом начнут атрофироваться, если совсем в тень попадут. Сразу замазывать раны антисептиком для ран на растениях - Садовым Варом или РанНет. Обсудить на форуме- 79 комментариев
-
- 1
-
-
- физиология растений
- агротехника
- (и ещё 1 )